



1. Background
Although Candida albicans is the most common etiological agent of candidiasis, non-albicans species have recently emerged among various systemic candidiasis forms (1). The most common non-albicans species include C. glabrata, C. tropicalis, C. krusei, and C. parapsilosis isolated from systemic and vulvovaginal candidiasis (2-4). Candiduria is one of the most common challenges in hospitalized patients, including both adults and children. Many studies have reported that non-albicans species may better adapt to the urinary tract (5). For example, Harris et al. believed that C. glabrata was well adapted to substrate availability, osmolality, and urine pH (6). Hence, the C. glabrata species complex has been introduced as the second agent of candiduria (7). According to the SENTRY Surveillance Program (2008 to 2009), C. glabrata was 13.7, 5.2, 15.7, and 23.5% prevalent in the Asia Pacific, Latin America, Europe, and North America, respectively (8).
Despite developing more typing methods, limited studies have been conducted on the typing of C. glabrata isolates, and few data are available on this species' genetic diversity (9). Techniques, such as electrophoretic karyotyping, multilocus sequence typing, randomly amplified polymorphic DNA, its typing, and microsatellite length polymorphism (MLP), have been introduced for C. glabrata typing in epidemiological investigation (10). Among these techniques, MLP is attractive for population structure analysis among microorganisms. Microsatellite length polymorphism is based on short tandem repeat regions that are highly polymorphic in microorganism genomes. Limited polymorphic loci have been introduced for C. glabrata typing, including MTI, ERG3, RPM2, GLM4, GLM5, and GLM6 (11). It is believed that multilocus analysis with these markers provides a rapid, reliable, and highly discriminatory tool for epidemiological investigation.
Data on antifungal susceptibility tests are essential to treating resistance cases of candidiasis. The C. glabrata species complex has shown reduced susceptibility to azole drugs, especially fluconazole (12). In a report by Pfaller et al., C. glabrata species from North America were resistant to anidulafungin, micafungin, and azoles at the rates of 3.2, 2.7, and 5.5 - 8.2%, respectively (8). The cross-resistance to echinocandins and azoles was reported in the C. glabrata species complex (13, 14). In contrast to azoles and echinocandins, polyenes, especially amphotericin B, are usually effective on the C. glabrata species complex (15, 16).
2. Objectives
We aimed to detect different genotypes of C. glabrata isolates using six microsatellite markers and the MLP technique. Our genotypes' association with other countries' genotypes was illustrated using a minimum spanning tree (MStree). We also investigated in vitro antifungal susceptibility and enzymatic activity profiles of the isolates.
3. Methods
3.1. Yeast Isolation and Identification
Candida glabrata isolates were collected from urine samples of pediatric patients admitted to the Abuzar Children's Hospital in Ahvaz, Iran, during the course of the study conducted in 2019. All the isolates were initially identified based on the phenotypic pattern on CHROM agar Candida media (CHROM agar Candida, France) and under microscopic examination. For accurate identification of isolates, genomic DNA was extracted and purified according to the method proposed by Looke et al. (17). DNAs were subjected to PCR amplification with V9G (5’-TTACGTCCCTGCCCTTTGTA-3’) and LS266 (5’-GCATTCCCAAACAACTCGACTC-3’) primers (18). Then, the Cardiogenic Department of the Rajaie Cardiovascular, Medical, and Research Center, Tehran, Iran, sequenced PCR products. Nucleotide sequences were edited and aligned with Mega 6 software and blasted in the GenBank database (NCBI) with a similarity of over 99%. Finally, sequence data were deposited in NCBI, and accession numbers were received.
3.2. Microsatellite Analysis
The isolates were genotyped using polymorphic microsatellite loci, including MTI, RPM2, ERG3, GLM4, GLM5, and GLM6. Forward primers of these markers were labeled with fluorochrome stains (FAM, HEX, and TAMRA). The multiplex reaction was performed according to the method developed by Abbes et al. (11). PCR products were analyzed using the capillary electrophoresis, and the fragment lengths were obtained using Gene Marker software (V3.0.0.0).
3.3. Antifungal Susceptibilities Tests
Antifungal susceptibility tests were carried out according to the CLSI M27 S4 guideline. Twenty-two C. glabrata isolates were tested against amphotericin B (Sigma - Aldrich, Germany), fluconazole (Serva, USA), itraconazole (Sigma), posaconazole (Sigma), voriconazole (Sigma), luliconazole (APIChem Technology, China), and caspofungin (Sigma). Finally, the minimum inhibitory concentration (MIC), MIC50, MIC90, and MIC geometric mean (GM) were calculated. Furthermore, CLSI M27 S4 (19) was used to interpret MIC values for fluconazole and caspofungin and the epidemiological cutoff value (ECV) (20) for amphotericin B, itraconazole, posaconazole, and voriconazole. Besides, C. albicans ATCC 10231 was used for quality control.
3.4. Enzymatic Activity
Specific substrates, including egg yolk, bovine serum albumin agar (Merck, Germany), tween 80 (Merck, Germany), and sheep blood (Bahar Afshan, Iran), were used for phospholipase, proteinase, esterase, and hemolysin activity, respectively, according to the study by Gharaghani et al. (18). Media were inoculated by the isolates and incubated at 37°C for one week for phospholipase, hemolysin, and proteinase activity. Esterase media were incubated at 25°C for ten days. Finally, the halo diameter and the colony diameter were measured, and the activity index of each enzyme was calculated. The enzymatic activity was interpreted based on the index presented by Price et al.: (1) 1 (negative), (2) 0.70 - 0.99 (low), (3) 0.40 - 0.69 (medium), and (4) 0.10 - 0.39 (high) (21).
3.5. Comparison of Genotypes from Different Countries
In the present study, the same microsatellite data on C. glabrata from China (22), Iran (23), Austria, Finland, Germany, Italy, Morocco, Netherlands, Nigeria, Portugal, South Africa, Spain, Greece, Turkey, UK, and Zimbabwe were extracted from published papers (24). The data were analyzed using BioNumeric Applied math software.
3.6. Statistical Analysis
Our study applied the UPGMA dendrogram to determine the genetic relationship between the C. glabrata isolates using BioNumeric Applied math software version 7.6. (https://www.applied-maths.com/quote-request?comm=1). Besides, the discriminatory power (DP) was calculated for each locus and a combination of six markers based on Simpson's index of diversity (http://insilico.ehu.es/mini_tools/discriminatory_power/index.php). Finally, the data were analyzed using Fstat software, and the Wright Fixation index (FST) was calculated (https://www.scienceopen.com/document?vid=b577cf2d-d522-4831-a4a7-d05d435e72cf).
4. Results
In the present study, we identified 22 (3.6%) C. glabrata isolates from 610 pediatric patients admitted to different wards, including seven (31.8%) nephrology wards, six (27.3%) neonatal intensive care wards, three (13.6%) infant wards, two (9.1%) pediatric wards, two (9.1%) pediatric mobile intensive care wards, one (4.5%) neurology ward, and one (4.5%) surgery ward. Of the patients, 13 (59.1%) and nine (40.9%) were female and male, respectively, with the age range > 1 year (n = 4, 18.2%), 1 - 3 years, (n = 8, 36.4%), 4 - 6 years (n = 5, 22.7%), 7 - 9 years (n = 3, 13.6%), and < 10 year (n = 2, 9.1%). The accession numbers were LC534380 - 85, LC534387 - 94, and LC567122 - 29.
4.1. Microsatellite Analysis
Our study detected 19 different multi-loci genotypes out of 22 C. glabrata isolates (G1 - G19). Among the 19 genotypes, G5 (n = 3, 15.8%) and G4 (n = 2, 10.5%) were the predominant genotypes. Generally, of the isolates, 22.7% had the same genotypes, whereas 77.3% had unique genotypes. Overall, we obtained 43 different alleles among six microsatellite loci, with GLM4 (n = 11) and ERG3 (n = 9) having the highest number of alleles, respectively (Table 1). The discriminatory power value varied from 0.76 for GLM5 to 0.91 for GLM4.
| Microsatellite Locus | Multiplex A | Multiplex B | ||||
|---|---|---|---|---|---|---|
| RPM2 | MTI | ERG | GLM4 | GLM5 | GLM6 | |
| Number of alleles | 5 | 5 | 9 | 11 | 6 | 6 |
| Index of diversity (DP) | 0.78 | 0.82 | 0.82 | 0.91 | 0.76 | 0.79 |
Also, the highest genetic diversity (haploid diversity) in the present study was obtained in a pediatric ward (H = 1.0, P < 0.05). It means that C. glabrata strains isolated from patients in the pediatric ward were genetically divergent from isolates obtained from patients in other wards (nephrology, neonatal intensive care, infant, pediatric mobile intensive care, neurology, and surgery wards). Moreover, the analysis of FST value between different wards showed that the FST index was lower than zero in all the cases. There was a slight difference between our genotypes with other countries. The minimum spanning tree was drawn based on differences in at least two alleles in the C. glabrata urinary isolates (Figure 1). Generally, five clonal clusters (CCs) were identified (CC1 to CC5), and isolates with identical genotypes were located in the same CC (CC5). Besides, except in CC5, there were no isolates with the same source in the other clusters.
. Dark, thin, and dashed lines related to one, two, or more different alleles, respectively, indicate the genetic distance among the genotypes.")
The minimum spanning tree showing the diversity of genotypes between different wards of the hospital. Each circle represents each genotype, and each halo surrounding each genotype represents clonal clusters (CCs). Dark, thin, and dashed lines related to one, two, or more different alleles, respectively, indicate the genetic distance among the genotypes.
4.2. Comparison of Genotypes of Candida glabrata from Different Countries
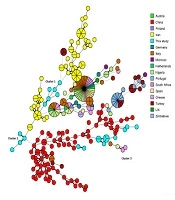
As shown in the MStree (Figure 2), two clusters of our genotypes (clusters 1 and 2) were related to the genotypes of C. glabrata isolates reported in a previous study in Iran, and the third cluster was completely connected with Chinese genotypes.

The minimum spanning tree representing the genotype diversity of Candida glabrata isolates in different countries. Each country is differentiated by color. Dark, thin, and dashed lines related to one, two, or more different alleles, respectively, indicate the genetic distance among the countries' genotypes.
4.3. Antifungal Susceptibility Profiles
The results of antifungal susceptibilities are demonstrated in Table 2. All the isolates were wild-type for amphotericin B, itraconazole, and voriconazole. In contrast, 77.3% of the isolates were the non-wild type for posaconazole. All the 22 C. glabrata isolates were sensitive to fluconazole, and resistance to caspofungin was observed in only three (13.6%) of the isolates. The lowest MIC range among antifungal drugs was related to luliconazole (0.0039 - 0.00048 μg/mL). Three (15.8%) of the MLP genotypes (G2, G11, and G18) were multi-resistance to posaconazole and caspofungin.
| Antifungals | Minimum Inhibitory Concentration, MIC (μg/mL) | Breakpoints, No. (%) | % ECV | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC Range | MIC50 | MIC90 | MICGM | S | DD | R | W | Non-W | |
| Amphotericin B | 0.0625 - 2 | 0.125 | 1 | 0.22 | - | - | - | 100 | 0 |
| Itraconazole | 0.125 - 2 | 0.5 | 2 | 0.19 | - | - | - | 100 | 0 |
| Voriconazole | 0.0078 - 0.00625 | 0.0312 | 0.0625 | 0.01 | - | - | - | 100 | 0 |
| Posaconazole | 0.0625 - 4 | 2 | 4 | 1.5 | - | - | - | 22.7 | 77.3 |
| Caspofungin | 0.0625 - 1 | 0.25 | 0.5 | 0.21 | 7 (31.8) | 12 (54.5) | 3 (13.6) | - | - |
| Fluconazole | 0.0312 - 16 | 4 | 8 | 2.42 | 22 (100) | - | - | - | - |
| Luliconazole | 0.00048 - 0.0039 | 0.00195 | 0.0039 | 0.0018 | ND | ND | ND | ND | ND |
Abbreviations: S, sensitive; DD, dose-dependent; R, resistance; ECV, epidemiological cutoff value; W, wild-type.
4.4. Enzymatic Secretion Pattern
The results of enzymatic secretion patterns in the urinary C. glabrata isolates are shown in Figure 3. As shown, 16 (72.7%) and 15 (68.2%) of the C. glabrata isolates had no phospholipase and esterase activities, respectively, and the other isolates had low activity for these enzymes. In comparison, the highest potency of enzymatic secretion was obtained in hemolysin 12 (54.5%) and proteinase 14 (66.5%) enzymes.
.")
The UPMGA dendrogram demonstrating the genotyping of the 22 urinary Candida glabrata isolates with antifungal susceptibilities profiles and enzymatic secretion patterns (W, wild-type; S, sensitive, R, resistance; DD, dose-dependent; N, negative; H, high activity; M, medium activity; and L, low activity).
5. Discussion
Candida glabrata is a crucial non-albicans species with increasing prevalence and resistance to antifungal drugs (1, 25). Evaluating the diversity of molecular genotypes with antifungal susceptibility profiles of C. glabrata is an essential technique for epidemiology investigation, population genetics analysis, and clinical practice guidelines. Some researchers used the six defined markers (ERG3, MTI, RPM2, GLM4, GLM5, and GLM6) for C. glabrata analysis and reported different discriminatory powers, including 0.964 (23), 0.941 (11), 0.88 (22), and 0.84 (26). However, in our study, the highest discriminatory power was calculated to be 0.978 for the six markers.
Nineteen multi-loci unrelated MLP genotypes were detected in the 22 C. glabrata isolates, including two (10.5%) identical genotypes (G4 and G5) and 17 (89.5%) singleton genotypes. However, different studies have reported various numbers of genotypes in the form of candidiasis. For example, in a previous study in Iran, 35 different genotypes, (39.3% singletons genotypes and 60.7% identical genotypes) were identified among 61 vaginal isolates of C. glabrata. (23). Abbes et al. (11) identified 37 genotypes for 85 C. glabrata isolates originating from different Tunisia sources. Of the isolates, 69.4% and 30.6% had shared and singleton genotypes, respectively. However, we observed no similarity between Abbes et al.’s genotypes detected in urine samples and our urine isolates. According to the MStree, two clusters of our genotypes (Clusters 1 and 2) were related to genotypes of C. glabrata isolates reported in a previous study in Iran (23). However, the third cluster was entirely connected with Chinese genotypes (22). In the present study, Fst was > 0.05, and according to Abbes et al.'s study, this species has a low genetic differentiation level in different hospital wards (27). In our hospital, 22.7% of the isolates had the same genotype. Moreover, Abbes et al. (9) observed 38% similarity in the genotypes and considered interpatient transfers of isolates in the hospital essential in transmitting diseases.
All the C. glabrata isolates were sensitive to fluconazole, and only three (13.6%) of the isolates were resistant to caspofungin. The highest resistance rate against fluconazole in the urinary C. glabrata isolates was 50% in Singla et al.'s report (28). In contrast, Patel and Bhavsar (29) reported that 20% of C. glabrata isolated from urine samples were resistant to fluconazole. Moreover, only one (5.6%) of urine isolates of C. glabrata had MIC > 64 µg/mL for fluconazole in Brazil (30). Hence, fluconazole is defined as the primary choice for candiduria treatment; however, amphotericin B can be used to treat azole-resistance isolates, C. glabrata, and C. krusei (28). Resistance in C. glabrata against caspofungin was diverse in reports, including 0.5, 4, 33.3, and 42.1% in different countries (13, 22, 31, 32), although several researchers indicated no resistance (33-35).
Our results indicated that all the C. glabrata isolates were wild-type for voriconazole, itraconazole, and amphotericin B. In a similar vein, Kooshki et al. (36) reported that C. glabrata isolates were all susceptible to amphotericin B, voriconazole, and itraconazole. Moreover, Nademi et al. (37) reported that C. glabrata isolated from nosocomial candiduria were all susceptible to amphotericin B at MIC ≤ 1, whereas only 25% of isolates in Singla et al.’s report (28) had resistance to amphotericin B. Furthermore, Patel and Bhavsar showed that C. glabrata isolates were all susceptible to voriconazole (29). Also, 13, 35.3, 50, and 77.8% of C. glabrata isolates were resistant to itraconazole in ST-Germain et al.’s (15), Sellami et al.’s (16), Singla et al.’s (28), and Badiee et al.’s (32) reports, respectively.
In the present study, 77.3% of the C. glabrata isolates were non-wild type (MIC > 1 µg/mL) for posaconazole (22). This result is inconsistent with the study conducted by Morales-Lo'pez et al. (38) and Zarei Mahmoudabadi et al. (31), showing that 100% of isolates were sensitive to posaconazole. However, Hou et al. (22) showed that only 7.3% of C. glabrata isolates from different sources were the non-wild type for posaconazole. Zaoutis et al. (33) reported a range of 0.5 to < 16 µg/mL with MIC90 = 2 µg/mL for 15 C. glabrata isolates. The deficient range of luliconazole, 0.00048 - 0.0039 µg/mL, with MIC90 = 0.0018 in our study is comparable to that in Taghipour et al.’s study (39). In our study, no differences were found regarding antifungal susceptibility or genotype profiles among the tested isolates associated with sources of isolates, similar to the results reported by Safdar et al. (40) and Abbes et al. (9).
Extracellular enzyme secretion was considered one of the virulence factors in Candida species. Our results indicated that C. glabrata isolates had no esterase and phospholipase activities. However, Sachin et al. (41) showed a low phospholipase and hemolytic activity rate (28.5 and 21.4%, respectively) in C. glabrata isolates. They also reported the highest enzymatic activity in proteinase (42.8%) among C. glabrata isolated from different sources. Deorukhkar and Saini (42) showed that enzymatic activity among C. glabrata isolates originated from various clinical specimens, varying from 15% for hemolysin activity to 38.3% in phospholipase production. They also reported a proteinase production rate of 36.6% in these isolates.
5.1. Conclusion
It was found that high genetic diversity existed among the urinary C. glabrata isolates. Moreover, based on the MStree, two clusters of our genotypes were related to C. glabrata genotypes in a previous study in Iran, and the third cluster was entirely connected with Chinese genotypes. The majority of the isolates were the non-wild type for posaconazole, but were rarely resistant to other antifungals. Furthermore, hemolysin and proteinase secreted as the main virulence factors among the urinary C. glabrata isolates.
