





1. Background
Streptococcus pneumoniae is a human bacterial pathogen considered as one of the main causes of community-acquired pneumonia, meningitis, and bacteremia and is a common cause of mortality worldwide. S. pneumoniae can colonize the upper respiratory tract including nasopharynx. The most common serotypes of S. Pneumoniae are 19F, 6B, 4, 14, 19A, 3, 23F, and 1. Bacterial attachment to host epithelial cells is required for the development of pneumococcal infections (1-4). Pneumococcus has several virulence factors which contribute to the colonization of the host mucosal surfaces (5). Neuraminidase is a virulence factor that cleaves sialic acid into glycoproteins, glycolipids, and oligosaccharides and contributes to pneumococcal pathogenesis in two ways: (1) decrease of the integrity of mucosa and facilitation of bacterial dissemination into lower layers; (2) facilitation of the attachment of surface receptors to host cells and bacterial colonization.
Two types of neuraminidase are encoded by nanA and nanB genes. Also, there is a homologous gene named nanC. The structural analysis of pneumococcal neuraminidases revealed that 50% of NanB and NanC sequences are similar, but 25% of their sequences are similar with that of NanA (6, 7). NanA is the primary virulence factor for the elimination of sialic acid in three types of neuraminidases. It also removes sialic acid from compounds such as lactoferrin and IgA2 protease (8).
NanA also targets the host glycogen molecules and mediates biofilm formation by pneumococci. nanA is upregulated during biofilm development (9). NanB cleaves sialic acid and provides carbon and energy sources for pneumococci growth. NanC function has remained unclear, but it is essential for invasive infections such as meningitis and hemolytic uremic syndrome (10-12). Since the available vaccines are expensive, serotype-dependent, and their immunogenicity is limited to serotypes included in the vaccine, scientists’ endeavors are concentrated on the development of a serotype-independent vaccine.
For this purpose, neuraminidases are studied in some countries, because this molecule has immunogenicity and is preserved in all strains (13). Serotyping of native and prevalent pneumococci in Iran, frequency of neuraminidases in these serotypes, and comparison of the results with those of other studies from other countries are necessary for obtaining more conclusive results for further investigations.
2. Objectives
The current study aimed at investigating the prevalence of pneumococcal nasopharyngeal carriage among unvaccinated children under six years of age in six provinces of Iran. The study also aimed at evaluating the frequency of pneumococcus serotypes and the neuraminidase virulence genes (nanA, nanB, and nanC) among the identified carriers.
3. Methods
The current cross-sectional study was conducted on 384 nasopharyngeal samples collected by sterile Dacron swabs from children under six years old referred to healthcare centers of six Iranian provinces including Tehran, Fars, Ardebil, Lorestan, Khorasan, and Sistan va Baluchistan from September 2013 to December 2016. The samples were then kept in a combined preserving medium (composed of beef extract 0.4 g, agar 1.2 g, casein 1.0 g, and NaCl 0.3 gin 100 mL double distilled water [BBL Co.]). The samples were sent to the Department of Bacteriology at Pasteur Institute of Iran for isolation and diagnosis. The participants were assigned to the three equal age groups of 2 - 24, 25 - 48, and 49 - 72 months. The nasopharyngeal swabs were streaked onto 5% sheep blood chocolate agar (Quelab Cat. No. 1601a) and then incubated at 37ºC in 5% CO2. The suspected pneumococcal colonies were subcultured onto chocolate agar.
3.1. Identification of Pneumococci
Pneumococci were identified based on the results of culture test and colony morphology, Gram staining, and biochemical tests including catalase, bile solubility and susceptibility of bacteria to optochin disks on chocolate agar plates (Mast diagnostics Cat. No. D42). Growth inhibition zones ≥ 14 mm in diameter around the optochin disks were considered as pneumococci. The primary detection of pneumococci was based on the appearance of the green zone of alpha-hemolysis surrounding colonies on chocolate agar, observation of lancet-shaped diplococci in Gram-stained smears and negative results in catalase test (14, 15).
The clearance of turbidity in tubes containing 2-McFarland bacterial suspension and sodium deoxycholate (bile salt) after 15 minutes was considered as pneumococci. The confirmatory method for the detection of pneumococci was based on molecular methods (16-19).
First, pneumococcal DNA was extracted and purified by Qiagen extraction kit (Cat No. 69525; Germany) and then cpsA gene was detected by PCR as the key test for confirmation. Specific primers for cpsA detection were provided as described in Table 1.
| Serotype | Primers (5’ - 3’) | Reference |
|---|---|---|
| 1 | F: CTC TAT AGA ATG GAG TAT ATA AAC TAT GGT TA | (20) |
| R: CCA AAG AAA ATA CTA ACA TTA TCA CAA TAT TGG C | ||
| 3 | F: ATG GTG TGA TTT CTC CTA GAT TGG AAA GTA G | (20) |
| R: CTT CTC CAA TTG CTT ACC AAG TGC AAT AAC G | ||
| 4 | F: CTG TTA CTT GTT CTG GAC TCT CGA TAA TTG G | (20) |
| R: GCC CAC TCC TGT TAA AAT CCT ACC CGC ATT G | ||
| 5 | F: ATA CCT ACA CAA CTT CTG ATT ATG CCT TTG TG | (20) |
| R: GCT CGA TAA ACA TAA TCA ATA TTT GAA AAA GTA TG | ||
| 6A/B | F: AAT TTG TAT TTT ATT CAT GCC TAT ATC TGG | (20) |
| R: TTA GCG GAG ATA ATT TAA AAT GAT GAC TA | ||
| 7F | F: CCT ACG GGA GGA TAT AAA ATT ATT TTT GAG | (20) |
| R: CAA ATA CAC CAC TAT AGG CTG TTG AGA CTA AC | ||
| 9V | F: CTT CGT TAG TTA AAA TTC TAA ATT TTT CTA AG | (20) |
| R: GTC CCA ATA CCA GTC CTT GCA ACA CAA G | ||
| 10 | F: GGT GTA GAT TTA CCA TTA GTG TCG GCA GAC | (20) |
| R: GAA TTT CTT CTT TAA GAT TCG GAT ATT TCT C | ||
| 11 | F: GGA CAT GTT CAG GTG ATT TCC CAA TAT AGT G | (20) |
| R: GAT TAT GAG TGT AAT TTA TTC CAA CTT CTC CC | ||
| 12F | F: GCA ACA AAC GGC GTG AAA GTA GTT G | (20) |
| R: CAA GAT GAA TAT CAC TAC CAA TAA CAA AAC | ||
| 14 | F: CTT GGC GCA GGT GTC AGA ATT CCC TCT AC | (20) |
| R: GCC AAA ATA CTG ACA AAG CTA GAA TAT AGC C | ||
| 15 | F: ATT AGT ACA GCT GCT GGA ATA TCT CTT C | (20) |
| R: GAT CTA GTG AAC GTA CTA TTC CAA AC | ||
| 16 | F: CTG TTC AGA TAG GCC ATT TAC AGC TTT AAA TC | (20) |
| R: CAT TCC TTT TGT ATA TAG TGC TAG TTC ATC C | ||
| 18C | F: CTT AAT AGC TCT CAT TAT TCT TTT TTT AAG CC | (20) |
| R: TTA TCT GTA AAC CAT ATC AGC ATC TGA AAC | ||
| 19F | F: GTT AAG ATT GCT GAT CGA TTA ATT GAT ATC C | (20) |
| R: GTA ATA TGT CTT TAG GGC GTT TAT GGC GAT AG | ||
| 19A | F: GTT AGT CCT GTT TTA GAT TTA TTT GGT GAT GT | (20) |
| R: GAG CAG TCA ATA AGA TGA GAC GAT AGT TAG | ||
| 22F | F: GAG TAT AGC CAG ATT ATG GCA GTT TTA TTG TC | (20) |
| R: CTC CAG CAC TTG CGC TGG AAA CAA CAG ACA AC | ||
| 23F | F: GTA ACA GTT GCT GTA GAG GGA ATT GGC TTT TC | (20) |
| R: CAC AAC ACC TAA CAC ACG ATG GCT ATA TGA TTC | ||
| 33F | F: GAA GGC AAT CAA TGT GAT TGT GTC GCG | (20) |
| R: CTT CAA AAT GAA GAT TAT AGT ACC CTT CTA C | ||
| 35B | F: GAT AAG TCT GTT GTG GAG ACT TAA AAA GAA TG | (20) |
| R: CTT TCC AGA TAA TTA CAG GTA TTC CTG AAG CAA G |
Polymerase Chain Reaction Primers for the Determination of the Main Pneumococcal Serotypes
3.2. PCR
The presence of neuraminidase genes in the isolated pneumococci was determined by the detection of cpsA gene using PCR. Using the primers provided for the detection of most prevalent serotypes of pneumococci (Table 2), all the confirmed isolates were then serotyped based on multiplex PCR (20).
| Genes | Primers | Size of the Gene (bp) | Reference |
|---|---|---|---|
| cpsA | F: GCA GTA CAG CAG TTT GTT GGA CTG ACC | 150 | (20) |
| R: GAA TAT TTT CAT TAT CAG TCC CAG TC | |||
| nanA | F: ATAGACGTGCGCAAAATACAGAATCA | 548 | (10) |
| R: GTCGAACTCCAAGCCAATAACTCCT | |||
| nanB | F: ACTAGGAGCTGTTAATCGTGAAGG | 492 | (10) |
| R: CCAATACCCGCAGGCATAACATC | |||
| nanC | F: TGGGGTAAGTACAAACAAGAA | 523 | (10) |
| R: CTAATGGTACTGGCGCAAAATCA |
Polymerase Chain Reaction Primers for the Detection of cpsA and Neuraminidase Genes
The multiplex PCR results were confirmed in the current study using standard strains including ATCC 6305, ATCC 6301, ATCC 49619, ATCC 49136, and ATCC 700677. Multiplex PCR was performed in a total volume of 25 µL as the reaction mixture containing 0.5 µL of dNTPs (10 mM), 0.5 µL of each primer (10 pmol), 2.5 µL of PCR buffer (10×), 1.5 µL of MgCl2 (25 mM), and 0.2 µL of Taq DNA polymerase (5U/µL) (Fermentas/thermofischer USA. Cat. No. EP0402). The PCR cycle was run at 94ºC for four minutes followed by 30 amplification cycles at 94ºC for 45 seconds, 56ºC for 45 seconds, and 72ºC for two minutes. PCR products were electrophoresed on 1% agarose gel at 100 V for one hour; then, they were stained with ethidium bromide and visualized under a UV transilluminator. The primers used for the detection of the three neuraminidase genes in the current study are shown in Table 3.
| Serotype | Provinces | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tehran (N = 260) | Lorestan (N = 260) | Ardabil (N = 260) | Khorasan (N = 260) | Fars (N = 260) | Sistan and Baluchestan (N= 260) | |||||||
| No. (%) | 95%CI | No. (%) | 95%CI | No. (%) | 95%CI | No. (%) | 95%CI | No. (%) | 95%CI | No. (%) | 95%CI | |
| 6A/B | 5 (1.9) | 0.6 - 4 | 3 (1.2) | 0.2 - 4 | 2 (0.8) | 0 - 3 | 1 (0.4) | 0 - 2 | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 |
| 14 | 5 (1.9) | 0.6 - 4 | 0 (0.0) | 0 - 2 | 2 (0.8) | 0 - 3 | 2 (0.8) | 0 - 3 | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 |
| 3 | 6 (2.3) | 0.9 - 5 | 1 (0.4) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 3 (1.2) | 0.2 - 4 | 0 (0.0) | 0 - 2 |
| 19A | 4 (1.5) | 0.4 - 4 | 4 (1.5) | 0.4 - 4 | 1 (0.4) | 0 - 2 | 1 (0.4) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 4 | 4 (1.5) | 0.4 - 4 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 |
| 19F | 3 (1.2) | 0.2 - 4 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 23F | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 2 (0.8) | 0 - 3 |
| 11 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 2 (0.8) | 0 - 3 |
| 18C | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 22F | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 9V | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 19 | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 12 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 33F | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 |
| 5 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| 12F | 1 (0.4) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 | 0 (0.0) | 0 - 2 |
| Non-typable | 6 (2.3) | 0.9 - 5 | 2 (0.8) | 0 - 3 | 1 (0.4) | 0 - 2 | 2 (0.8) | 0 - 3 | 0 (0.0) | 0 - 2 | 2 (0.8) | 0 - 3 |
| Total | 43 (2.8) | 2 - 3.7 | 10 (0.6) | 0.3 - 1.2 | 9 (0.6) | 0.3 - 1.1 | 11 (0.7) | 0.4 - 1.3 | 9 (0.6) | 0.3 - 1.1 | 8 (0.5) | 0.2 - 1.0 |
Frequency of Positive Pneumococcus Serotypes in the Six Provinces of Iran
S. pneumoniae ATCC49610 was used as positive control in PCR. Polymerase chain reaction in the present study was performed in a total volume of 25 μL with 1X PCR buffer in 2.5 μL, 0.2 mM of dNTPs, 1 U of Taq DNA polymerase, 1.5 mM of MgCl2, 2.5 μL of each primer, and 1μL of DNA template. Thermocycling was performed in Eppendorf Gradient Mastercycler apparatus with the following steps: 98ºC for 30 seconds (denaturation), 64ºC for 30 seconds (annealing) for nanA and 55ºC for 30 seconds for nanB and nanC, 72ºC for 60 seconds (extension) and final extension at 72ºC for five minutes. Polymerase chain reaction product was electrophoresed on 1% agarose gel at 100 V for one hour. The final PCR products were stained with ethidium bromide and then illustrated by UV light. The DNA bands were analyzed by comparison of the visualized bands with a 100-bp ladder.
To confirm the results of PCR for nanA, nanB, and nanC genes in the current study, two positive PCR products were randomly selected and sequenced with direct Sanger sequencing (Biomatik Co. Canada).
3.3. Statistical Analysis
Data were collected, revised and analyzed by SPSS software (version 16). Frequencies of identified carriers, pneumococcus serotypes, and neuraminidase genes were expressed as number and percentage. The Wilson score interval was used to estimate 95% confidence interval (CI) for the key outcomes (frequencies). Difference in the frequency of pneumococcal nasopharyngeal carriage among six provinces of Iran was determined by the chi-square (χ2) test. P-value less than 0.05 was considered significant.
4. Results
Of the 384 collected swabs, 119 (31%) and 92 (24%) samples were confirmed as pneumococci by phenotypic and genotypic (detection of cpsA gene) assays, respectively (Figure 1). Of the 92 genetically-confirmed pneumococci, the serotype of 79 (86%) isolates was determined using PCR. The most frequent serotype in the studied samples was 6A/B with the frequency of 14.1% (95% CI: 8.5, 22.7%). The least frequent serotypes were 12F and 5, with the frequency of 1.1% each (95% CI: 0.2, 5.9%). The frequency of the identified pneumococcal serotypes in different provinces and overall is presented in Tables 3 and 4, respectively (21).
. The lanes 2 - 14 represent suspect alpha-hemolytic streptococci that lanes 6 - 14 were detected as Pneumococci because these lanes indicate the <i>cpsA</i> and the lanes 3 - 5 removed from our study because they were not detected as Pneumococci.")
Gel electrophoresis for cpsA gene as confirmatory test of Pneumococci isolates. M. indicates the 100 bp. ladder marker, lane 1: Standard strain of Pneumococcus for cpsA. (ATCC49610). The lanes 2 - 14 represent suspect alpha-hemolytic streptococci that lanes 6 - 14 were detected as Pneumococci because these lanes indicate the cpsA and the lanes 3 - 5 removed from our study because they were not detected as Pneumococci.
| Serotypes | Frequency of Serotypesa | Frequency of Neuraminidases Genesb | ||||||
|---|---|---|---|---|---|---|---|---|
| No. (%) | 95% CIc | nanA | nanB | nanC | ||||
| No. (%) | 95% CIc | No. (%) | 95% CIc | No. (%) | 95% CIc | |||
| 6A/B | 13 (14.1) | 8.5, 22.7 | 13 (100) | 77.2, 100 | 12 (92.3) | 66.7, 98.6 | 9 (69.2) | 42.4, 87.3 |
| 14 | 11 (12.1) | 6.8, 20.2 | 11 (100) | 74.1, 100 | 9 (81.9) | 52.3, 94.9 | 7 (63.7) | 35.4, 84.8 |
| 3 | 11 (12.1) | 6.8, 20.2 | 11 (100) | 74.1, 100 | 8 (72.8) | 43.4, 90.3 | 7 (63.7) | 35.4, 84.8 |
| 19A | 11 (12.1) | 6.8, 20.2 | 11 (100) | 74.1, 100 | 11 (100) | 74.1, 100 | 9 (81.9) | 52.3, 94.9 |
| 4 | 5 (5.4) | 2.3, 12.1 | 5 (100) | 56.6, 100 | 4 (80) | 37.6, 96.4 | 4 (80) | 37.6, 96.4 |
| 19F | 5 (5.4) | 2.3, 12.1 | 5 (100) | 56.6, 100 | 3 (60) | 23.1, 88.2 | 3 (60) | 23.1, 88.2 |
| 23F | 5 (5.4) | 2.3, 12.1 | 5 (100) | 56.6, 100 | 4 (80) | 37.6, 96.4 | 1 (20) | 36.2, 62.4 |
| 11 | 4 (4.3) | 1.7, 10.7 | 4 (100) | 51.0, 100 | 4 (100) | 51.0, 100 | 1 (25) | 4.6, 69.9 |
| 18C | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 2 (100) | 34.2, 100 | 1 (50) | 9.5, 90.5 |
| 22F | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 2 (100) | 34.2, 100 | 2 (100) | 34.2, 100 |
| 9V | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 2 (100) | 34.2, 100 | 1 (50) | 9.5, 90.5 |
| 19 | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 1 (50) | 9.5, 90.5 | 1 (50) | 9.5, 90.5 |
| 12 | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 2 (100) | 34.2, 100 | 0 (0) | 0, 65.8 |
| 33F | 2 (2.1) | 0.6, 7.6 | 2 (100) | 34.2, 100 | 1 (50) | 9.5, 90.5 | 1 (50) | 9.5, 90.5 |
| 5 | 1 (1.1) | 0.2, 5.9 | 1 (100) | 20.7, 100 | 1 (100) | 20.7, 100 | 1 (100) | 20.7, 100 |
| 12F | 1 (1.1) | 0.2, 5.9 | 1 (100) | 20.7, 100 | 1 (100) | 20.7, 100 | 1 (100) | 20.7, 100 |
| Non serotypes | 13 (14.1) | 8.5, 22.7 | 13 (100) | 77.2, 100 | 12 (92.4) | 66.7, 98.6 | 8 (61.5) | 35.5, 82.3 |
| Total | 92 (5.9) | 4.8, 7.2 | 92 (100) | 96.0, 100 | 79 (85.9) | 77.3, 91.6 | 57 (62.0) | 51.8, 71.2 |
Frequency of Pneumococcal Serotypes and Neuraminidases Genes in a Sample of Iranian Children Under Six Years of Age (N = 92a)
The presence of neuraminidase genes was determined in the 92 genetically-confirmed isolates (Figure 2). All the confirmed isolates harbored the nanA gene, but nanB and nanC were only detected in 85.9% and 62.0% of the isolates, respectively. Also, 87% of the samples collectively harbored nanA and nanB, 62% concomitantly carried nanA and nanC, and 55% harbored nanB, nanC, and nanA genes. Furthermore, 13% and 38% of all isolated strains did not carry nanB and nanC genes, respectively. Table 4 illustrates detailed data on the frequency of different serotypes.
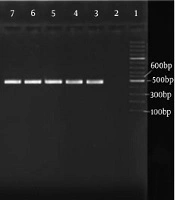
 Electerophoresis gel of <i>nanA</i> gene PCR. Lane 1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 4 - 12 represent test isolates. (<i>nanA</i> gene size: 548 bp). (B) Electerophoresis gel of <i>nanB</i> PCR. Lane 1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 4 - 10: represent test samples, lane 8: negative for <i>nanB</i>. (<i>nanB </i>gene size: 492 bp). (C) Electerophoresis gel of <i>nanC</i> PCR. Lane1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 6, 10 and 11 were negative samples.")
(A) Electerophoresis gel of nanA gene PCR. Lane 1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 4 - 12 represent test isolates. (nanA gene size: 548 bp). (B) Electerophoresis gel of nanB PCR. Lane 1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 4 - 10: represent test samples, lane 8: negative for nanB. (nanB gene size: 492 bp). (C) Electerophoresis gel of nanC PCR. Lane1: 100-bp ladder, lane 2: negative control, lane 3: positive control, lanes 6, 10 and 11 were negative samples.
5. Discussion
Pneumococcal infections are increasing after the emergence of non-vaccine serotypes, capsule switching, and replacement of new serotypes among different communities (22). There are two types of vaccines (i.e., conjugate and polysaccharide) to prevent pneumococcal infections with each covering partial and limited groups of serotypes (23-25). Different serotypes have distinct geographical distributions. Pneumococci express several types of antigens and virulence factors that help them cause infections in humans. One of these antigens is neuraminidase enzyme (Nan) that hydrolyses the neuraminic acids on human cell surfaces. Pneumococci express three variants of neuraminidase. Several pneumococcal proteins and antigens are selected as vaccine candidates including neuraminidases.
The presence of Nan variants, NanA, NanB and NanC, varies among serotypes. Determination of the frequency of Nan variants in native serotypes is important for vaccine studies; thus, the prevalence of different serotypes and nan genes was investigated in the current research. Four serotypes (i.e., 19A, 6, 3, 23F) of S. pneumoniae accounted for 55.7% of all strains isolated from both nasal carriage and clinical samples in Tehran. Serotype 19A was the most frequent one in the two groups of samples (21, 26).
Vaccine serotypes used in conventional vaccines cover only the American serotypes and is ineffective for other geographical serotypes (27). Since the production of conjugated vaccines is expensive, there is a great limitation for the usage of all serotypes in these vaccines. They are produced in limited numbers from polysaccharides (28). To develop an effective vaccine and eliminate remarked conflicts, it is recommended to focus on protein-based extending vaccines as ideal alternatives for conventional vaccines. Thus, the virulence factors of pneumococcus, except the capsule, could be promising vaccine candidates.
As mentioned earlier, neuraminidases are pneumococcal proteins that play key roles in bacterial colonization in the early stages of infection and invasion to organs such as the central nervous system and the incidence of hemolytic uremic syndrome in infected patients (29-31). Thus, determination of the prevalence of neuraminidase genes in Iranian strains is of great importance. The prevalence of neuraminidase genes and its relationship with pneumococcal serotypes in native strains as the main objectives of the current study were determined by PCR.
Serotyping results of the current study were compared with those of other studies performed worldwide in order to explore differences and similarities; for example, Pai et al. (20) perfectly studied isolates of pneumococci by multiplex PCR, and the comparison of the results with those of the current study showed that pneumococcal serotypes were more prevalent in Iran. It can be concluded that the numbers of isolates, nanA, nanB, nanC, and serotypes according to the sampling site are comparable and the analysis of the results indicates that nanA and nanB distribution was not different among distinct serotypes, but results indicated that the presence of nanC was higher in invasive serotypes.
The investigation of pneumococcal virulence factors can provide more information as to its pathogenesis and lead to the development of more appropriate vaccine candidates for the prevention of pneumococcal diseases. Proper vaccines based on neuraminidase proteins (i.e., NanA, NanB and NanC) as pneumococcal virulence factors can result in more immunity, cost-saving and resolutions for some failures in developing pneumococcal vaccines. The prevalence of neuraminidase genes in clinical isolates and expression of these genes can also be evaluated for obtaining more information.
5.1. Conclusion
The analysis of the current study results indicated a correlation between pneumococcal serotypes and the prevalence of nanA and nanB genes; nanA gene was detected in 100%, nanB in 85.9%, and nanC in 62% of the isolated strains. Although the serotypes 4 and 12F were the most frequent ones in strains isolated from invasive infections, only one strain identified as serotype 12F harbored three neuraminidase genes. The prevalence rate of nanC gene in some serotypes found in the current study including 19A, 6A/B, 19F, and 22F was respectively 81.9%, 69.2%, 60%, and 100%. Some other serotypes such as 18C, 5, 12F, 19, and 19V were scarce, but could be the causes of pneumococcal invasive infections. Since a limited number of serotypes were identified in the current study, for certainty about lack of correlation between pneumococcal serotypes and neuraminidase genes, conducting further extensive studies is recommended.
