





1. Background
2. Objectives
3. Methods
3.1. Physical and Chemical Properties and Posttranslational Patterns and Motifs
| Server | URL | Predicted Localization |
|---|---|---|
| CELLO | http://cello.life.nctu.edu.tw/ | Extracellular, cytoplasmic |
| WoLF | PSORT http://wolfpsort.org/ | Extracellular |
| PSORT | II http://psort.hgc.jp/form2.html | Cytoplasmic, nuclear |
| SLPFA | http://sunflower.kuicr.kyoto-u.ac.jp/~tamura/slpfa.html | Extracellular |
| SLP-Local | http://sunflower.kuicr.kyoto-u.ac.jp/~smatsuda/slplocal.html | Extracellular |
| SherLoc2 | http://abi.inf.uni-tuebingen.de/Services/SherLoc2 | Extracellular |
3.2. Prediction of RNA Secondary Structure
3.3. Prediction of Secondary and Tertiary Structures
3.4. Prediction of T and B Cell Epitopes for SRS3 Protein and Comparison of Antigenicity with Applied Genes in Vaccines
4. Results
4.1. Physical and Chemical Characteristics Study of tSRS3 Using Protparam Program
4.2. Prediction of Modification Sites
4.3. Prediction of Trans-Membrane Structure and Sub-Cellular Localization

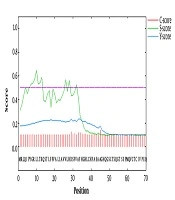
Signal peptide predicted within the SRS3 sequence. The C-score is high at the position immediately after the cleavage site. The S-score distinguishes between positions within signal peptides from those of the mature part of the proteins. The Y-score is a geometric average of the C-score and the slope of the S-score.
4.4. Phylogenetic Analysis of SRS3

Evolutionary relationship between representative SRS family members from the life cycle
4.5. Prediction of Secondary and Tertiary Structures of tSRS3
, extended strand (Ee), beta turn (Tt), and random coil (Cc)")
The SRS3 secondary structure prediction of alpha helix (Hh), extended strand (Ee), beta turn (Tt), and random coil (Cc)
 were modeled with 100.0% confidence by the single highest scoring template (BSR4), a, α-helix; b, random coil; c, β-folded; B, ligand-binding site results for the SRS3 protein. Blue colors list amino acid residues observed in cluster 10 of the predicted binding site; C, ProSA-web z-score SRS3 protein plot. The z-score indicates overall model quality. The Z-scores of SRS3 protein chain A in PDB were determined by X-ray crystallography (Pale blue) or NMR spectroscopy (bold blue) concerning their lengths. The plot shows results with a z-score of ≤ 10. The z-score of SRS3 is -5.7 and is shown as a large dot. The value of -5.7 is in the range of native conformations.")
A, The tertiary structure prediction of SRS3 by Phyre2; 324 residues (98.7% of SRS3 sequences) were modeled with 100.0% confidence by the single highest scoring template (BSR4), a, α-helix; b, random coil; c, β-folded; B, ligand-binding site results for the SRS3 protein. Blue colors list amino acid residues observed in cluster 10 of the predicted binding site; C, ProSA-web z-score SRS3 protein plot. The z-score indicates overall model quality. The Z-scores of SRS3 protein chain A in PDB were determined by X-ray crystallography (Pale blue) or NMR spectroscopy (bold blue) concerning their lengths. The plot shows results with a z-score of ≤ 10. The z-score of SRS3 is -5.7 and is shown as a large dot. The value of -5.7 is in the range of native conformations.
4.6. In Silico Prediction of T Cell Epitopes for SRS3 Antigen
| MHC-I Allele | Position | Peptide Sequences | Server | Vaxijen Scores |
|---|---|---|---|---|
| HLA-A*1101 | 167 - 175 | IVGCTQEGK | BIMAS | 1.7863 |
| HLA-A*1101 | 167 - 175 | IVGCTQEGK | ProPred-I | |
| HLA-A*1101 | 167 - 175 | IVGCTQEGK | EpiJen | |
| HLA-A*1101 | 247 - 256 | LQENCDEKY | RANKPEP | 0.985 |
| HLA-A*1101 | 269 - 277 | VSQAGTSNK | Consensus (ann/smm) | 0.5653 |
| HLA-A*1101 | 246 - 256 | PLQENCDEK | SVM | 1.1607 |
| HLA-A*1101 | 144 - 153 | DSQTVTCAY | NetCTL1.2 | 0.864 |
| HLA-A*0201 | 15 - 24 | STLFFVALLA | Consensus | 1.6329 |
| HLA-A*0201 | 16 - 23 | LFFVALLA | SVMHC | 2.0760 |
| HLA-A*0201 | 112 - 121 | GMTDPQSCV | BIMAS | 1.2069 |
| HLA-A*0201 | 179 - 188 | CLLHVTLSA | EpiJen | 0.8877 |
aAntigenicity of full-length protein epitopes was calculated using VaxiJen.
| HLA-A | Antigen | Peptide Sequences | Position | Vaxijen Scores |
|---|---|---|---|---|
| HLA-A*1101 | SAG1 | KSFKDILPK* | 224 - 232 | 0.4842 |
| HLA-A*1101 | GRA6 | AMLTAFFLR* | 164 - 172 | 1.9220 |
| HLA-A*1101 | GRA7 | RSFKDLLKK* | 134 - 142 | -0.2503 |
| HLA-A*1101 | SPA | SSAYVFSVK* | 250 - 258 | 0.8154 |
| HLA-A*1101 | SAG2C | STFWPCLLR* | 13 - 21 | 0.8148 |
| HLA-A*1101 | SAG2D | ALLEHVFIEK | 63 - 72 | 0.9153 |
| HLA-A*1101 | BSR4 | TTRFVEIFPK | 284 - 293 | 1.0332 |
| HLA-A*1101 | SAG3 | VVGHVTLNK | 80 - 88 | -0.4365 |
| HLA-A*1101 | SRS3 | IVGCTQEGK** | 167 - 175 | 1.7863 |
| HLA-A*0201 | GRA6 | FMGVLVNSL* | 29 - 37 | 1.9980 |
| HLA-A*0201 | SPA | GLAAAVVAV* | 82 - 90 | 0.5290 |
| HLA-A*0201 | GRA3 | FLVPFVVFL* | 25 - 33 | 2.2250 |
| HLA-A*0201 | SAG2C | FLSLSLLVI* | 38 - 46 | 0.7125 |
| HLA-A*0201 | SAG2D | FMIAFISCFA* | 180 - 189 | 1.1592 |
| HLA-A*0201 | SAG2x | FMIVSISLV* | 351 - 359 | 0.1095 |
| HLA-A*0201 | SAG3 | FLTDYIPGA* | 136 - 144 | 1.4166 |
| HLA-A*0201 | SRS3 | LFFVALLA** | 16 - 23 | 2.0760 |
aCandidate epitopes were used in the vaccine * and the elected epitope of SRS3 protein **. Antigenicity of full-length protein epitopes was calculated using VaxiJen.
4.7. In Silico Prediction of B Cell Epitopes for SRS3 Antigen and Evaluation of Antigenicity
| B Cell Epitope | Prediction Server | Amino Acid Positions | Scores | Vaxijen Scores |
|---|---|---|---|---|
| HRVTVSPKDPTFTLDCSSDG | BCPred | 210 | 0.998 | 1.2979 |
| VCPAGMTDPQSCVSPSGTIP | BCPred | 108 | 0.975 | 0.9927 |
| GSCTSQGTSSPMQVCTCIVP | BCPred | 49 | 0.955 | 0.8694 |
| MQPASFTTSYCPA | BCPred | 231 | 0.879 | 0.7269 |
| TGPLQENCDEKYASIL | ABCpred | 244 | 0.72 | 1.3467 |
| TLSARASSKDSQTVTC | ABCpred | 184 | 0.74 | 1.3257 |
| SLLSGDGASAVAWVGD | ABCpred | 123 | 0.69 | 1.1767 |
| PEAQKKVLVGCKASTA | ABCpred | 288 | 0.72 | 1.0681 |
| GETSNHAVVVSAEKNQ | ABCpred | 71 | 0.87 | 0.9510 |
| ELTITSSSPSSAWFLS | ABCpred | 313 | 0.73 | 0.9448 |
| GTIPFASLLSGDGASA | ABCpred | 124 | 0.77 | 0.8582 |
| TLDCSSD | BAP | 222 | High | 1.7363 |
| WWQVSQA | BAP | 266 | High | 1.6518 |
| NHAVVVSAE | BAP | 75 | High | 1.2996 |
| QKKVLVGCKASTA | BAP | 291 | High | 1.1761 |
| ASAVAWVGDE | BAP | 130 | High | 1.1352 |
| PQSCVSPSGTIPFASLLSG | BAP | 116 | High | 0.9466 |
| HFAEAAGHQGSCTSQGTSSPM | BepiPred | 40 | High | 1.084 |
| SLLSGDGASAVAWVGDEVN | BepiPred | 130 | High | 1.046 |
aThe server assessment threshold is 0.5.
bNon-antigen.
cHigh score = High antigenicity.
