


1. Background
Menopause is a normal period that accounts for one-third of a woman’s life during which, hormonal changes cause permanent cessation of menstruation (1). The lack of estrogen in women after menopause creates oxidative stress by releasing free radicals and reactive oxygen species, causing some metabolic diseases and imposing the risk of various cardiovascular diseases such as high blood pressure, decreased bone density, and premature cell aging (2). There is ample evidence that menopause is associated with increased oxidative stress and increased reactive oxygen species, leading to mutations in the mitochondrial genome and subsequent cell death through necrosis or apoptosis (3). Cell death is the final stage of cell damage (4). Researchers believe that apoptosis increases with age and menopause (5), and Bax, as an important in the process of apoptosis, plays a special role in cognitive disorders in this mechanism (6). By increasing oxidative stress, the tumor suppressor protein inhibits muscle cell proliferation and accelerates cell death by activating the apoptotic process and caspase 9 (7).
Exercise affects many aspects of brain function and has a wide range of effects on overall brain health so that it has shown to have excellent benefits on learning and protection against neurodegeneration and reduction of depression, especially in postmenopausal women (8). Researchers have shown that exercise training with reducing oxidative stress, increasing neurotrophins, inhibiting apoptotic proteins such as Bax, increasing Bcl-2, and inhibiting inflammatory factors can improve cognitive function in rats (9, 10). However, the type and intensity of exercise training have different effects on neurological and cognitive adaptations in rats. Moderate-intensity endurance training can improve cognitive memory, and resistance training has more favorable effects on spatial memory. Exercise training reduces oxidative stress, but resistance training also increases the levels of nitrite and free radicals (9).
Nowadays, along with performing exercises, it is proposed to use medicinal plants with different biological properties due to fewer side effects and treatment costs. Saffron, scientifically known as Crocus sativus, belongs to the Iris family, which contains a compound called crocin. Crocin has antioxidant properties and the ability to destroy free radicals, and can also significantly reduce oxidative damage in various tissues (11). In previous studies, researchers have well demonstrated the anti-apoptotic and antioxidant role of crocin, as crocin consumption could improve DNA repair markers in rodents’ brains by reducing oxidative stress markers such as thiobarbituric acid reactive substances (TBARS) and nitrite levels mechanism in the hippocampus (12). Daily consumption of 25 mg/kg crocin peritoneally augmented Bcl-2 gene expression and declined Bax gene expression in rats (13). Researchers suggested that taking 0.5 ml of crocin reduces Bax and increases Bcl-2 and antioxidant enzymes in rats, which decreases apoptosis by reducing lipid peroxidation (14).
Estrogen reduction conditions in humans can be modeled by ovariectomy in young rats. This animal model is suitable for the study of the hypoestrogenic effect on adipocytes because of weight and abdominal obesity increases in rats with the removal of ovaries (ovariectomy) (15). Regular exercise and physical activity can reduce the percentage of body fat and body mass index in postmenopausal women aged 50 - 59 (16). Mohammad ibn Zakaria Razi believes that the consumption of saffron causes anorexia, and it is suitable for preventing overweight (17). Daily consumption of at least 5 mg/kg crocin for two months could significantly reduce the weight of obese rats. Therefore, there are positive effects for exercise training and crocin intake on the reduction of body weight and Bax gene expression while these two factors increase during menopause. On the other hand, there is inadequate research on the concurrent effect of aerobic training and crocin intake during menopause.
2. Objectives
The present research investigated the effect of eight weeks of aerobic training and crocin intake on the weight and Bax gene expression of ovariectomized rats.
3. Methods
3.1. Subjects
An experimental study was conducted with 45 eight-week-old female rats weighing approximately 180 to 200 g. Rats were purchased from the Reproduction Center and Animal House of Marvdasht. They went through a seven-day adaptation period in standard conditions with an ambient temperature of 22°C to 27°C, a relative humidity of 50%, and controlled light. They were given water and food ad libitum during the research period, and their food was prepared from the Pars Livestock Food Company (Tehran, Iran).
The animals were anesthetized with an intraperitoneal injection of 50 mg/kg ketamine solution and 4 mg/kg xylazine, and then the abdominal area was shaved. After rinsing with the betadine solution, the sides of the abdomen were split between breasts 2 and 3 next to the thigh muscle, and the ovary was separated and removed by a cutter device (18). The animals were kept under controlled conditions for one month to create osteoporosis (19). The weight of rats after this period reached 210 to 230 g.
Rats were then randomly divided into five groups of nine subjects, including (1) control (C); (2) sham (Sh); (3) crocin (Cr); (4) aerobic training (AT); and (5) aerobic training + crocin intake (AT + Cr). Rats in groups 4 and 5 performed training for three sessions per week for eight weeks. Rats in groups 5 and 3 intraperitoneally received 25 mg/kg body weight of crocin per day (Sigma Aldrich, Germany) dissolved in normal saline (20). To control the effects of the injection on the research variables, the sham group received the crocin solvent intraperitoneally each day. Twenty-four hours after the last training session at the end of week 8, the rats went through surgery to measure the parameters studied. They were then anesthetized with xylazine 2% (10 mg/kg body weight) and ketamine 10% (50 mg/kg body weight) after about five minutes. The head was cut by surgery, and then the brain and hippocampus were removed by specialists and kept at -70°C until further tests.
3.2. Aerobic Training Protocol
Aerobic training was performed as five sessions per week of incremental running on a rodent treadmill for eight weeks. In this way, the running activity started at a speed of 15 m/min on a treadmill for 15 min on a zero-degree inclination in the first session and continued in such a way that in the last four weeks, the activity speed reached 26 m/min, the duration of each training session increased to 60 min, and the inclination of the treadmill increased to 10 degrees. In addition, about 5 min of warming up and 5 min of cooling down were devoted to each set of training (21). The warming and cooling program was implemented at a speed of 8 m/min and an inclination of zero degrees.
3.3. Measurement of Bax Gene Expression
For molecular studies at the gene expression level, RNA was first extracted from the hippocampal tissue according to the manufacturer's protocol (Sinagen, Iran). Then, using the light absorption property at a wavelength of 260 nm, the concentration and purity of the RNA sample were obtained quantitatively using the following equation.
C (µg/µL) = A260 × ε × d/1000
After extracting RNA with very high purity and concentration from all the studied samples, the cDNA synthesis steps were performed according to the manufacturer's protocol and then, the synthesized cDNA was used to perform the reverse transcription reaction. The designed primers were first examined for genes, and then the expression of genes was investigated using the quantitative (q)RT PCR method (Table 1).
| Gene | Sequence of Primer | Product Size, bp |
|---|---|---|
| Bax | F: 5’GCAGACGGCAACTTCAACTG3’ | 174 |
| R: 5’TGTCCAGCCCATGATGGTTC3’ | ||
| B2m | Forward: 5’- CGTGCTTGCCATTCAGAAA -3’ | 244 |
| Reverse: 5’-ATATACATCGGTCTCGGTGG -3’ |
The Sequence of Research Primers of Genes in Real-Time Polymerase Chain Reaction
3.4. Statistical Analysis
To attain the results, data were reported using descriptive statistics as mean and standard deviation. Then, the Shapiro-Wilk test was used to investigate the normal distribution of the data. To compare the study groups, one-way Analysis of Variance (ANOVA) and Tukey’s post hoc test were used. All statistical calculations were run using SPSS (version 24), and a significant level of P < 0.05 was considered.
4. Results
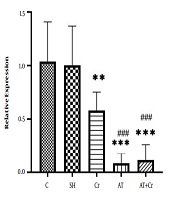
The results of one-way ANOVA showed that eight weeks of aerobic training and crocin intake (P = 0.001, F = 31.34) had a significant effect on reducing Bax gene expression in the brain hippocampal tissue of ovariectomized rats. The results of Tukey’s post hoc test showed that Bax gene expression did not differ significantly between the C and Sh groups (P = 0.98). However, Bax gene expression was significantly lower in the Cr (P = 0.01), AT (P = 0.001), and AT + S (P = 0.001) groups than in the C and Sh groups. Also, Bax gene expression was significantly lower in the AT (P = 0.003) and AT + Cr (P = 0.024) groups than in the Cr group, and no significant difference was observed between the AT + Cr group and the AT group (P = 0.94) (Figure 1).
The results of one-way ANOVA showed that eight weeks of aerobic training and crocin intake made a significant difference (P = 0.001, F = 404.85) in the weight of ovariectomized rats. The results of Tukey’s post hoc test showed that the weights did not differ significantly in the C and Sh groups (P = 0.98). However, the weights showed to be significantly lower in the Cr (P = 0.001), AT (P = 0.001), and AT + Cr (P = 0.001) groups than in the control and sham groups. Also, the weights were significantly lower in the AT (P = 0.001) and AT + Cr (P = 0.001) groups than in the Cr group. There was no significant difference between the AT + Cr group (P = 0.054) and the AT group (Figure 2).
 and **, P ≥ 0.01, significantly lower than in the Sh and C groups; ###, P = 0.001 significantly lower than in the Cr group.")
Levels of Bax gene expression in the brain hippocampal tissue of rats in the five study groups. ***, P = 0.001) and **, P ≥ 0.01, significantly lower than in the Sh and C groups; ###, P = 0.001 significantly lower than in the Cr group.
 from the C and Sh groups; ¥, significantly different (P = 0.001) from the Cr group.")
The weight of rats in the five study groups. *, Significant decreases in the mean scores of the pretest-posttest; #, significant increases in the mean scores of the pretest-posttest; £, significantly different (P = 0.001) from the C and Sh groups; ¥, significantly different (P = 0.001) from the Cr group.
5. Discussion
The results of the current study revealed that eight weeks of aerobic training reduced Bax gene expression in the hippocampal tissue of ovariectomized rats. The results of this study also showed that aerobic training significantly reduced apoptosis caused by ovariectomy in the hippocampus region of the brain. In line with this finding, Kim et al. (22) reported the beneficial effects of endurance training on reducing gyrus apoptosis. As one of the possible mechanisms for the protection of neurons, exercise can increase the capacity to block free radicals (23). Aksu et al. (24) showed that physical activity increases the levels of antioxidant enzymes in different parts of the brain, which, in turn, increases the antioxidant capacity of the brain. Exercise can block the formation of free radicals (25). On the other hand, inconsistent with the present study, one bout of long-term aerobic training reduced the levels of Bcl-2 to Bax ratio protein immediately after training in the muscle tissue of rats (26). The results of another study showed that two months of voluntary activity had no meaningful effect on Bcl-2 and Bax in rats (27). One possible reason for the inconsistency is the differences in the statistical population and examined tissue.
The results of the present study showed that two months of crocin intake had a meaningful effect on the reduction of Bax gene expression in the hippocampal tissue of ovariectomized rats. Crocin can protect saturated fatty acids in cell membranes by reducing free radicals from cyclophosphamide. It has also been reported that crocin, through the inhibition of oxidative stress and the reduction of lipid peroxidation, prevents reactive oxygen species to increase and thus inhibits caspases and prevents apoptosis (28). Regarding the effect of crocin on Bax, some researchers reported that receiving 0.5 ml of crocin reduced Bax and increased Bcl-2 and antioxidant enzymes (14). Also, Moradi et al. (13) reported that the consumption of 25 mg/kg crocin peritoneally per day for eight weeks had a meaningful effect on reducing Bax gene expression in diabetic rats. The consumption of crocin through the free radical scavenging mechanism and the increased expression of transcription proteins from antioxidant enzymes (29) can synergistically increase antioxidant enzymes in ovariectomized rats. The above studies observed a reduction in oxidative stress and anti-apoptotic effects, which may be due to the molecular mechanism of saffron and its anti-inflammatory properties.
Another study found that two months of aerobic training combined with the intake of crocin could reduce Bax gene expression in the hippocampal tissue of ovariectomized rats. Reactive oxygen species can increase the expression of antioxidant enzymes and other cellular protein protectors to adapt to oxidative stress and maintain cellular homeostasis by activating dual intracellular pathways. As mentioned earlier, crocin may inhibit the production of reactive oxygen species by reducing lipid peroxidation, thereby inhibiting caspases and preventing the induction of apoptosis through reducing Bax gene expression. Exercise also blocks apoptosis pathways by increasing the expression and activity of kinase B protein, phosphorylation of Bcl-2 family anti-apoptotic proteins, and inactivation of progressive apoptotic proteins such as Bax, or through the direct inhibition of caspase activity (25). Most importantly, exercise and the simultaneous use of crocin in both interventions could reduce Bax and anti-apoptosis by inhibiting caspase activity and reducing reactive oxygen species (13).
Research has shown that eight weeks of aerobic training reduced the weight of ovariectomized rats. Also, ovariectomy stimulated adipocyte hypertrophy and increased the levels of the epidermal growth factor. Besides, these factors were effective in stimulating obesity in ovariectomized rats (30). In this study, the weight of rats significantly reduced after eight weeks of aerobic training compared to the ovariectomized control group, which is consistent with a study by Vilacxa Alves et al. (31), who observed similar changes. Regular aerobic training increases the gene expression of lipolysis enzymes, beta-oxidation, Krebs and electron transport chains, enhances mitochondrial density, and leads to fat consumption instead of carbohydrate use for energy production; thus, it reduces body fat and causes weight loss and body mass index reduction (32). On the other hand, inconsistent with the present study, Pourrahim et al. (33) did not observe a significant difference in the weight of experimental and control groups of postmenopausal women. The reason for inconsistency can be differences in the statistical population (humans versus animals) and different training protocols.
In this study, crocin consumption significantly reduced the weight of ovariectomized rats compared to the control group. There are a few studies that have examined the effects of crocin intake on ovariectomized rats, so the results are discussed based on analogous research in this regard. Consistent with the results of the present study, Kianbakht et al. (34) showed that crocin intake reduced the weight of the subjects under study. Pure saffron contains flavonoid compounds with abundant antioxidant properties. The most important carotenoids in the saffron extract are crocin and safranal (35). It is possible that by controlling oxygen free radicals and eliminating associated metabolic disorders, it is effective in reducing body fat and blood. One of the possible mechanisms of the crocin effect is that it prevents pancreatic lipase activity and reduces fat mass and increases insulin sensitivity, as mentioned in previous studies (34).
The results of other research showed that eight weeks of aerobic training combined with crocin intake reduced the weight of ovariectomized rats. In a similar study, Rajabi et al. (36) investigated the effect of a period of aerobic training and oral consumption of sprout powder (400 mg daily for two months) in obese diabetic women and showed a significant reduction in weight and the percentage of fat and an improvement in the fat profile compared to the control group and aerobic training group. Meamarbashi and Rajabi (37) showed that saffron supplementation had a significant effect on the oxidation of fat and carbohydrates, so it could be suggested that saffron supplementation increased the role of fat in energy production during exercise.
Due to the antioxidant role of crocin and the modulating effects of exercise training on the balance of oxidative-antioxidative stress in brain tissue, one of the limitations of the present study seems to be the lack of the study of variables related to oxidative-antioxidative stress. It is suggested that these variables be examined in future studies. It also appears that the Bax variable alone does not indicate apoptotic changes, so further studies are proposed to investigate further markers along with apoptotic assessment methods such as Tunnel and H & E.
5.1. Conclusions
It seems that aerobic training and crocin consumption, alone and synergistically, can reduce apoptosis and decrease the weight of ovariectomized rats. According to the findings of the present study, aerobic training combined with crocin intake can reduce the weight of ovariectomized rats. Also, aerobic training and the concurrent use of crocin and aerobic training can reduce Bax gene expression in the hippocampal tissue of ovariectomized rats.
