



1. Backgrounds
Dexamethasone (DEX), a synthetic glucocorticoid, is increasingly used as a doping agent by athletes. It is also used as symptomatic treatment for various diseases, such as asthma, osteoarthritis, and transplant rejection. Furthermore, it is a potent anti-inflammatory, immunosuppressive, and powerful painkiller (1). Chorionic usage of DEX increases generation of intracellular reactive oxygen species (ROS). DEX, in turn, can cause metabolic and reproductive disorders (2).
Studies have shown that dyslipidemia, hyperglycemia, insulin resistance, increased lipid peroxidation, impaired testosterone production, spermatogenesis, and sperm parameters are the main adverse reproductive side effects of DEX (2). In addition, it has been reported that DEX leads to testicular tissue damage by affecting the gonadal axis of the anterior pituitary gland and testicles. Hashemitabar et al. reported this effect mainly through the induction of apoptosis by the Fas/FasL signaling pathway in Leydig and germ cells (3). Previous studies have shown that a decrease in testosterone levels is associated with an increase in visceral fat and insulin resistance (4). The chronic consumption of DEX can cause oxidative stress related to ROS and lipid peroxidation and activation of inflammatory cytokines. The role of oxidative stress in causing reproductive disorders and its progress towards infertility has been well demonstrated, which can be due to the changes in the hormonal disorders such as reduction of gonadotropins and testosterone, sperm parameters, and testicular tissue (5). Nrf2 is a major transcription agent in controlling many aspects of cellular homeostasis in response to oxidative changes (6). Furthermore, Nrf2 plays a noted role in regulating glucose metabolism through the transcription of ingredients of the pentose phosphate pathway (7). Studies in which pharmacological interventions have been used as Nrf2 activators have produced significant results. First, the increase of Nrf2 target enzymes such as superoxide dismutase (SOD) and oxygenase-1 (HO-1) can improve insulin signaling and glucose consumption in conditions of insulin resistance (8). Second, it inhibits oxidative stress associated with inflammation by activating Nrf2, which also exacerbates insulin resistance (8). Third, the activation of Nrf2 by activating the mitogen-activated protein kinase (MAPK) signaling pathway can inhibit obesity and its complications (9). Therefore, the activation of transcription factors such as Nrf2 is an important component in signaling cascades related to oxidative stress responses.
In recent years, researchers have well shown that regular exercise can increase the activity of the antioxidant defense system due to adaptation to oxidative stress (10). On the other hand, sedentary lifestyle has negative effects on human health and can lead to a significant decrease in Nrf2 expression and activity. These effects are mainly due to the disorder in the signaling pathway of the Nrf2/keap-1 complex, which leads to an increase in the intracellular amount of ROS and, as a result, the damage caused by them (11). The results of studies on animal models have demonstrated that exercise plays a key role in improving the redox state of cells, and increases the activity and transcription of Nrf2. Adaptation to endurance training can increase Nrf2 activation, improve respiratory capacity, and increase ATP production during exercise (12). According to the results from animal and human studies, exercise can reduce oxidative stress and strengthen the antioxidant system. Several studies have investigated the effect of aerobic exercise on antioxidant status and markers of oxidative stress (13). The intensity and duration of exercise play important roles in strengthening the antioxidant system during exercise by affecting the ROS production. Some available evidence shows that the production of free radicals increases after intense exercise. However, it is believed that regular exercises improve the antioxidant status of the body. Insufficient research has been done on the effect of regular exercise with different intensities on testicular tissue in an animal model treated with dexamethasone.
Physical activity can enhance fertility (14). A period of aerobic exercise can improve sperm quality and fertility (15). In addition to the role of sports activities in improving the antioxidant defense system, which can be an important factor in controlling diseases such as obesity, insulin resistance, and other metabolic diseases, using plant-based antioxidants has attracted the attention of many researchers in the last decade. In the meantime, the safflower plant with the Persian name Kafishe has wide therapeutic applications in traditional medicine. It has been used as a therapeutic agent to enhance the reproductive function (16). In a study on laboratory mice by Modaresi, it was reported that safflower extract significantly increased the concentration of serum testosterone, and may have been a relevant factor contributing to changing male reproductive potential and affecting the endocrine function of the testicles (17). In another study by Nasiri et al. on male rats, it was discovered that safflower seed extract had the potential to improve the quality of sperm produced in rats with type 2 diabetes (18). Considering these results, it seems that regular physical activity with medium to high intensity as well as the consumption of safflower seed, can reduce the oxidative stress in the testicular tissue and improve fertility. In addition, no study has ever explored the effect of two types of moderate and high intensity endurance training along with safflower seed extract on testicular function in rats consuming dexamethasone.
2. Objectives
This study, therefore, aimed to determine whether endurance training with high and moderate intensity with the consumption of safflower extract had a beneficial effect on the expression of the NRf2 gene, total antioxidant capacity, MDA, and testosterone in testicular tissue of rats consuming dexamethasone.
3. Methods
3.1. Animals and Study Design
In this study, 45 adult male Wistar rats (aged 16 - 14 weeks) with an average weight of 283 ± 17 were obtained from Pasteur Institute of Iran and transferred to the laboratory. All stages of the study were approved by the ethics committee in biological research affiliated to Mazandaran University with the ethical code IRMU.IREC.1398.007. The animals were kept in standard lab conditions. All animals had free access to water and rat chow. After a week of acclimatization to the laboratory environment, the animals were divided into nine groups as fallow: (1) control; (2) dexamethasone; (3) dexamethasone + safflower extract; (4) dexamethasone + intense exercise training; (5) dexamethasone + moderate exercise training; (6) dexamethasone + intense exercise training + safflower extract; (7) dexamethasone + moderate exercise training + safflower extract; (8) intense exercise training + safflower extract; and (9) moderate exercise training + safflower extract. Subcutaneous injection of dexamethasone with a dose of 8 mg/kg was performed for five days (19).
3.2. Training Protocols
To familiarize the animals with the process of the training protocol, the training groups were allowed to run on the treadmill for 10 minutes at a speed of 10 to 15 meters per minute for three days a week. After the injection of dexamethasone, the training groups were allowed to perform two endurance training protocols with moderate intensity (Table 1) and high intensity (Table 2) with equal distances traveled for ten weeks. Moderate and high-intensity exercises were performed from 15 m/min for 10 minutes and 20 m/min for 8 minutes, respectively, and continued by the end of the eighth week. They were increased gradually at the same distances until the intensities of 28 m/min for 60 minutes and 34 m/min for 50 minutes were reached. The intensity and duration of the exercises remained unchanged up to the end of the tenth week.
In all training sessions, 5 minutes of warming up and cooling down at a speed of 10 meters per minute were performed.
| Week | First | Second | Third | Fourth | Fifth | Sixth | Seventh | Eighth | Ninth | Tenth |
|---|---|---|---|---|---|---|---|---|---|---|
| Speed (m/min) | 15 | 18 | 20 | 22 | 23 | 24 | 24 | 28 | 28 | 28 |
| Duration (min) | 10 | 20 | 30 | 30 | 40 | 40 | 50 | 60 | 60 | 60 |
Moderate Intensity Endurance Training Protocol
| Week | First | Second | Third | Fourth | Fifth | Sixth | Seventh | Eighth | Ninth | Tenth |
|---|---|---|---|---|---|---|---|---|---|---|
| Speed (m/min) | 20 | 22 | 24 | 26 | 28 | 30 | 31 | 32 | 32 | 32 |
| Duration (min) | 8 | 16 | 24 | 28 | 32 | 36 | 40 | 60 | 60 | 60 |
High Intensity Endurance Training Protocol
3.3. Extraction Method and How to Use Safflower Seed Extract
Extraction was performed based on the method described by Nimrouzi et al. (20). Briefly, the process of grinding safflower seeds was completed at room temperature. To this end, 250 g of seed powder was mixed with 70% ethanol in an Erlenmeyer flask, and shaken with a shaker for one hour. Afterwards, the mixture was filtered using a filter paper. The obtained liquid was placed in a rotary, so that the solvent evaporated at a temperature of 40°C. The residue was used as safflower seed extract. The preparation of the extract dosage (500 mg/kg of body weight) was also done using normal saline, and it was applied to the desired groups through daily gavage at a specific hour before the initiation of exercise (20).
3.4. Sampling and Measuring Variables
Forty-eight hours after the last training session, the rats were anesthetized with an intraperitoneal injection of a combination of ketamine (50 mg/kg) and xylazine (5 mg/kg) in order to eliminate the acute effect of training after a night of fasting. Blood samples were taken from the inferior vena cava, and the testicular tissues was taken for molecular, biochemical, and histological evaluation; then it was washed with cool normal saline, placed in microtubes, frozen using liquid nitrogen immediately, and stored at -80°C. The blood samples obtained from the animals were kept for 10 minutes under laboratory conditions in a 37°C incubator and then centrifuged for 15 minutes at 3500 rpm. The serum was separated from the clot, and the samples were stored at -20°C for measuring biochemical parameters. The tissue testosterone level was measured by an ELISA kit provided by Zelbio (Germany). The levels of MDA and TAC were measured by ELISA method via Zelbio commercial kit (Germany).
3.5. Gene Expression
Total RNA was extracted from rat testis tissue using Denazist kit. The quality and quantity of extracted RNA was evaluated by agarose gel electrophoresis and UV spectrophotometry (Nanodrop Iranian, Iran). Then the extracted RNAs were treated using DNasI, RNase-free enzyme. Synthesis of cDNA was performed by using the Yekta Tehiz-Azma kit (Iran) based on the company’s instructions. Oligo-dT primer was used to make cDNA. Specific primers for the amplification of Nrf2 gene were designed using Primer Premier 5 software, and, following the sequence of primers, they were synthesized by Bioneer (South Korea). The sequence of reverse primers for this gene is shown in Table 3. Polymerase chain reaction of cDNA samples for Nrf2 gene was performed using Cybergreen kit from Ampliqon (Denmark) in Rotor gene Corbett 6000 machine. Normalization of the target gene data was practiced against HPRT1 gene as the housekeeping gene. The desired gene expression was calculated by adopting the Livak method (2-ΔΔCT).
| Gene Sequence | Access Number | Product Length (Open Pair) |
|---|---|---|
| Nrf2 | NM_031789.2 | 104 |
| F 5´- GCTGCCATTAGTCAGTCGCTCTC-3´ | ||
| R 5´- ACCGTGCCTTCAGTGTGCTTC -3´ | ||
| HPRT1 | NM_012583.2 | 123 |
| F 5´-TTCTTTGCTGACCTGCTGGAT -3´ | ||
| R 5´-TATGTCCCCCGTTGACTGGT-3´ |
Specifications of the Primers Used in the Real-time PCR Process
3.6. Statistical Analysis Method
Graphpad Prism software was used to record the data, and one-way analysis of variance tests was performed to analyze them. The results were shown as mean ± standard deviation. Kolmogorov-Smirnov (KS) test was used to determine the normality of data distribution. The equality of variances was measured by Levene’s test. The LSD post hoc test was performed to determine the significant difference among the groups. A significance level of P ≥ 0.05 was considered.
4. Results
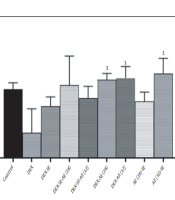
The results of Nrf2 gene expression in the studied groups are shown in Figure 1. The results showed that the testicular expression of Nrf2 gene decreased in the dexamethasone group although this decrease was not significant compared to that in the control group. The expression of Nrf2 gene in the dexamethasone-extract moderate-intensity exercise, dexamethasone-extract high-intensity exercise, and dexamethasone high-intensity exercise groups increased significantly compared to that in the dexamethasone group (Figure 1).
")
Nrf2 gene expression changes in the studied groups ($ Significant difference compared to dexamethasone group)
The results revealed that the MDA level increased significantly in the dexamethasone group compared to that in the control group (Figure 2). However, its level decreased in the dexamethasone groups treated with extract, moderate, and intense exercise training compared to that in the dexamethasone group. The largest decrease was observed in the dexamethasone- extract high-intensity group (Figure 2).
")
MDA Changes in the studied groups; * Significant difference compared to the healthy control group; $ Significant difference compared to dexamethasone group; # Significant difference compared to dexamethasone-SE group; ‡ Significant difference with dexamethasone-SE-AE group (32)
The results also demonstrated that although the testicular level of TAC decreased in the dexamethasone group, this decrease was not significant compared to that in the control group. However, its level significantly increased in the dexamethasone moderate-intensity exercise and dexamethasone high-intensity exercise groups compared to that in the dexamethasone group (Figure 3).
")
TAC changes in the studied group ($ Significant difference compared to dexamethasone group)
Testosterone level changes in testicular tissue are shown in Figure 4. Testosterone levels in the dexamethasone group did not change significantly compared to that in the control group. The testosterone levels significantly increased in the dexamethasone-extract high-intensity exercise training, dexamethasone moderate-intensity exercise training, and extract high-intensity exercise training groups compared to those in the dexamethasone groups. Testosterone levels significantly increased in the dexamethasone-extract high-intensity exercise training group compared to those in the dexamethasone-extract, dexamethasone high-intensity exercise training, and moderate-intensity exercise training extract groups. Testosterone levels significantly increased in the dexamethasone high-intensity exercise training group compared to those in the dexamethasone-extract moderate-intensity exercise training, dexamethasone moderate-intensity exercise training, and high-intensity exercise training extract groups. In the extract-exercise training with moderate intensity group, moreover, a significant decrease was detected compared to that in the dexamethasone-exercise training with moderate intensity and extract-exercise training with intense intensity groups (Figure 4).
; ¥ Significant difference with dexamethasone-AE group (32); ‡ Significant difference with dexamethasone-SE-AE group (32); ^ Significant difference with AE (28)-SE group; † Significant difference with dexamethasone-AE group (28)")
Testosterone changes in the studied groups; * Significant difference compared to the healthy control group; $ Significant difference compared to DEX group; # Significant difference compared to dexamethasone-SE group; & Significant difference compared to the dexamethasone-SE-AE group (28); ¥ Significant difference with dexamethasone-AE group (32); ‡ Significant difference with dexamethasone-SE-AE group (32); ^ Significant difference with AE (28)-SE group; † Significant difference with dexamethasone-AE group (28)
5. Discussion
Our study results showed that the injection of dexamethasone in rats led to oxidative damage by increasing MDA and decreasing the total antioxidant capacity. Moreover, dexamethasone had the potential to reduce the expression of Nrf2 as a controlling factor of oxidative status and the activity of antioxidant enzymes such as SOD and CAT. However, it failed to reduce tissue testosterone levels. It has been well-documented that the reduction of antioxidant capacity leads to infertility (21). Sadeghzadeh et al. showed that dexamethasone led to apoptosis and reduction of the average number of spermatocytes and spermatids (1). Dexamethasone can induce cell apoptosis (22), and increase the formation of free radicals (1). Our results, in line with these findings, also revealed that DEX suppressed the Nrf2 gene in testicular cells and reduced testosterone biosynthesis. Dexamethasone can prevent luteinizing hormone (LH) (1, 23), which was consistent with these results. In the present study, the level of MDA in testis in the DEX group was significantly higher than that in the control group, which was consistent with the results from previous studies (24-26). In our study, it was also found that dexamethasone decreased the serum level of testosterone, which was in agreement with the results from previous studies (27), suggesting that dexamethasone significantly reduced the transcription of genes encoding enzymes responsible for testosterone biosynthesis such as P450SCC (chain cleavage p450 side) (1).
In our study, safflower seed oil decreased MDA level, increased antioxidant capacity, and showed a non-significant decrease in Nrf2 gene expression compared to the dexamethasone group. Moreover, testosterone in this group may not have been significantly affected by safflower extract compared to that in the DEX or control groups. Adequate levels of cellular respiration factor and antioxidant levels are essential for the activity of the hypothalamus-pituitary-gonadal axis (18). Nasiri et al. investigated the effect of safflower on the activity of the hypothalamus-pituitary-gonadal axis, and reported that the levels of gonadotropins in rats suffering from type 2 diabetes were impaired, which may have been one of the factors contributing to the reduction of testosterone production in these patients (18). However, they showed that the consumption of safflower seed oil had the potential to improve the status of gonadotropins, inhibit oxidative damage, and strengthen the antioxidant system in testicular tissues and, therefore, improve testosterone level (18). It has been reported that safflower seed oil and its extract can improve oxidative damage and zinc homeostasis in rats with metabolic syndrome (20). Therefore, it is reasonably assumed that safflower seed oil can increase the expression of Nrf2 and the activity of the hypothalamus-pituitary-gonadal axis by reducing the cell-destructive factors and thus increasing the testosterone of the testicular tissue (28).
In this study, the effect of ten weeks of moderate and high intensity exercise training on oxidative indices and testosterone level was also investigated. It was observed that ten weeks of moderate and high intensity exercise in the dexamethasone groups decreased the level of MDA and increased the level of Nrf2. These findings were consistent with those form the study by Arslan et al. (29). In our study, the intensities of exercise training were compared and it was found that the high-intensity training was more effective than the low-intensity training in reducing MDA, although the difference was not statistically significant. However, our results were contrary to the findings of Xi et al. who observed that as the intensity of exercise increased, MDA also increased (30). The inconsistency may have been due to the difference in the type of disease since Xi et al. investigated rats with osteoarthritis (30). In our study, a noticeable increase was also observed in the amount of MDA factor in high intensity but not in low intensity compared to the dexamethasone + safflower group. This may have been attributed to the inherent property of high-intensity exercise, compared to low-intensity exercise, that increased MDA.
In this study, it was detected that exercise training increased Nrf2 expression and total antioxidant capacity compared to the consumption of safflower seed extract, although this change was not statistically significant. However, it has been reported that high-intensity exercise, not moderate-intensity, is able to reduce the cellular respiration factor in dexamethasone-treated rats (31). Our findings were consistent with the findings from the study by Baar et al. who showed that exercise may have reduced the number of cellular respiration factors (31). Increasing the total antioxidant capacity and Nrf2 in dexamethasone consuming mice with exercise training can constrain the risk factors related to infertility. Similar to our study, the study by Jannatifar et al. found that the expression of Nrf2 prevented the increase of oxidative stress, and had the potential to improve the qualitative parameters of semen in men with asthenoteratozoospermia (32). Studies on animal and human models have shown that oxidative damage is associated with hypothalamus-pituitary-testis axis activity disorder through the decreased secretion of LH and FSH in diabetic men. In our study, therefore, it was found that exercise training with saffron was associated with improving oxidative damage and increasing testosterone levels, which may have been due to the improved LH and FSH levels. Our findings were in line with the results from the study by Bonen et al. who reported that heavy exercise training induced significant increases in sex hormones in untrained subjects, whereas such increases did not occur in trained subjects exercising at the same absolute workload (33). It is worth mentioning that our study was the first one to report the effects of exercise training with safflower extract on the secretion of sex hormones. On the other hand, the increase in anti-inflammatory factors and antioxidant capacity caused by exercise increases testosterone production, which can be effective in controlling diseases related to infertility (34). In our study, it was observed that high-intensity exercise training with safflower extract significantly increased the testosterone levels compared to the dexamethasone and dexamethasone + safflower extract groups, which may have been attributable to the increased stimulation leading to the release of anabolic and steroid hormones due to exercise training. In this study, low-intensity exercise training was found capable of exerting a significant effect on body hormones. Exercise training along with safflower oil supplement can increase testosterone capacity. This increase can be partially attributed to the increase in antioxidant capacity and decrease in cell-destroying factors in animals consuming dexamethasone. Moreover, the most encouraging result was obtained in the combined group of high-intensity exercise with safflower extract, which had a synergistic effect.
