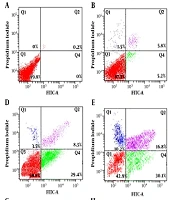
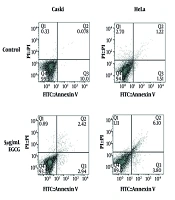
The current study concentrated on antitumoral effects of digoxin on HeLa cell line and showed that digoxin decreases the proliferation competency of cells. It also suggested that apoptosis and senescence induction might be the contributing mechanisms to digoxin function.
Digoxin, a cardiac glycoside, is used commonly (
13). It is an inexpensive and well-tolerated drug used widely as a treatment for heart failure and arrhythmia. Digoxin is still one of the most frequently used medicines in spite of a number of recently developed therapeutic agents for the treatment of heart failure, and remains the first choice of treatment (
14).
Reduction of cancer prevalence in patients with cardiac problems who used digoxin recorded in epidemiological studies have suggested the anticancer capability of Na
+/K
+ ATPase inhibitors in such cancers as prostate, breast, lung, or leukemia (
14).
Our findings revealed that digoxin can decrease viability and proliferation of HeLa cell line, meaning that it has antitumoral effects. It is suggested that cardiac glycosides increase the cytosolic Ca
2+ concentration and this mechanism explains the antitumor effects of these pharmacological agents in cancer patients (
15). In several experimental models, intracellular or extracellular Ca
2+ chelators, Ca
2+ channel blockers, and calmodulin antagonists are able to delay or decrease apoptosis. Damage of intracellular Ca
2+ homeostasis through inhibition of the Na
+, and K
+- ATPase by cardiac glycosides induce apoptosis in different cell types including tumor cell lines (
16).
Some forms of cardiac glycosides in clinically relevant doses could inhibit proliferation and induce apoptosis in prostate cancer cells. These findings suggest that the regulation of intracellular Ca
2+ concentration may induce or increase the apoptotic induction in human cancer cells and improve a new target in therapeutic strategies in cancer chemotherapy (
17).
Additionally, it is discovered that cardiac glycosides and their derivatives are strong potential antiproliferative agents, tumor-specific, and exert their cytotoxic effects through apoptosis (
18,
19).
It has been found that glycosides have the ability to inhibit the activity of Na
+, K
+-ATPase and lead to intracellular Ca
2+ increase (
20,
21). Disturbance in regulation of these ions activates a series of intracellular pathways that in turn changes cellular structure or gene expression. A variety of hydrolytic enzymes such as proteases, nucleases, and lipases activate an increase in intracellular Ca
2+. This process accounts for effectors of Ca
2+ elicited toxicity. Therefore, these changes may have a pivotal role in cellular toxicity (
17).
It is believed that gene expression patterns change during cancer development. Hypoxia-inducible Factor 1 (HIF-1) controls oxygen delivery through angiogenesis and metabolic modification to hypoxia via glycolysis in human cancers. Normally, there is hypoxia in tumors, and HIF-1α is overexpressed as a result of intratumoral hypoxia. The findings obtained from biopsies of brain, breast, cervical, esophageal, oropharyngeal and ovarian cancers showed a correlation between HIF-1α overexpression with treatment failure and mortality. In other words, increase in HIF-1 activity accelerates tumor progression and its inhibition could be considered a novel approach in cancer therapy (
22,
23).
It has been proved that risk of uterus cancer incidence in women who recently took digoxin significantly increased compared with those patients who did not receive digoxin. However, no change was obtained in cervical cancer risk (
24).
Although it has been proved that some glycosides have antitumoral effects, our knowledge is limited, and thus more studies should be conducted to clarify their mechanisms in cell toxicity and molecular field.





.")