



1. Background
2. Objectives
3. Methods
3.1. Identification and Editing of Cytochrome Oxidase I Gene Sequences
3.2. Phylogeny and Determining the Evolutionary Path
3.3. Molecular Diversity Indices
3.3.1. Molecular Diversity Indices
3.3.1.1. Polymorphic Sites
3.3.1.2. Haplotypes and Haplotype Frequency
3.3.1.3. Nucleotide Diversity (π)
3.3.1.4. Total Number of Mutations
3.3.1.5. Nucleotide Difference Between Populations
3.3.1.6. Conservation Indices
| No. | Scientific Name | ID Number in the NCBI Database |
|---|---|---|
| 1 | Apis andreniformis Aan 4.1 | LC427579.1 |
| 2 | Apis andreniformis voucher AAT95 | MT670338.1 |
| 3 | Apis andreniformis voucher AAT97 | MT670340.1 |
| 4 | Apis cerana isolate 8 | OK253002.1 |
| 5 | Apis cerana indica voucher ACIOR1 | OP168349.1 |
| 6 | Apis cerana indica breed Honey bee | MW093739.1 |
| 7 | Apis koschevnikovi Ak1_TB | LC461206.1 |
| 8 | Apis koschevnikovi Ak1_HST | LC461204.1 |
| 9 | Apis koschevnikovi Ak1_BL | LC461205.1 |
| 10 | Apis florea voucher UBN3 | OK350423.1 |
| 11 | Apis florea voucher UBN1 | OK350421.1 |
| 12 | Apis florea voucher SMI9 | OK350397.1 |
| 13 | Apis dorsata isolate Jatoli | OQ346377.1 |
| 14 | Apis laboriosa voucher ALM6 | MT679379.1 |
| 15 | Apis laboriosa voucher ALM5 | MT679378.1 |
| 16 | Apis laboriosa voucher ALM1 | MT679374.1 |
| 17 | Apis nigrocincta AnBK4.2 | LC534595.1 |
| 18 | Apis nigrocincta AnBK3.3 | LC534593.1 |
| 19 | Apis nigrocincta AnBK3.2 | LC534592.1 |
| 20 | Apis mellifera voucher HY42 | OP435367.1 |
| 21 | Apis mellifera voucher HY41 | OP435365.1 |
| 22 | Apis mellifera isolate mellifera | ON680902.1 |
| 23 | Apis mellifera isolate ACKU | OM320635.1 |
| 24 | Apis mellifera meda isolate DUHOK29-9 | OP020350.1 |
| 25 | Apis mellifera meda isolate DUHOK29-7 | OP020348.1 |
| 26 | Apis mellifera meda isolate DUHOK29-6 | OP020347.1 |
4. Results
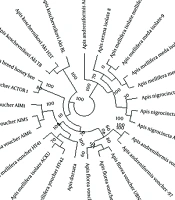
4.1. Experiment and Analysis of Genetic Distance and Phylogenetic Tree
| Variables | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Specie 1 | |||||||||||||||||||||||||
| Specie 2 | 1.293 | ||||||||||||||||||||||||
| Specie 3 | 1.293 | 0.000 | |||||||||||||||||||||||
| Specie 4 | 0.119 | 1.712 | 1.712 | ||||||||||||||||||||||
| Specie 5 | 1.221 | 0.124 | 0.124 | 1.168 | |||||||||||||||||||||
| Specie 6 | 1.221 | 0.129 | 0.129 | 1.482 | 0.013 | ||||||||||||||||||||
| Specie 7 | 1.173 | 0.123 | 0.123 | 1.302 | 0.114 | 0.098 | |||||||||||||||||||
| Specie 8 | 1.173 | 0.123 | 0.123 | 1.302 | 0.114 | 0.098 | 0.000 | ||||||||||||||||||
| Specie 9 | 1.162 | 0.119 | 0.119 | 1.285 | 0.111 | 0.098 | 0.003 | 0.003 | |||||||||||||||||
| Specie 10 | 1.372 | 0.096 | 0.096 | 1.483 | 0.108 | 0.119 | 0.114 | 0.114 | 0.111 | ||||||||||||||||
| Specie 11 | 1.372 | 0.096 | 0.096 | 1.483 | 0.108 | 0.119 | 0.114 | 0.114 | 0.111 | 0.000 | |||||||||||||||
| Specie 12 | 1.370 | 0.094 | 0.094 | 1.480 | 0.105 | 0.117 | 0.111 | 0.111 | 0.108 | 0.002 | 0.002 | ||||||||||||||
| Specie 13 | 1.306 | 0.120 | 0.120 | 1.390 | 0.117 | 0.103 | 0.124 | 0.124 | 0.127 | 0.134 | 0.134 | 0.132 | |||||||||||||
| Specie 14 | 1.233 | 0.115 | 0.115 | 1.560 | 0.100 | 0.090 | 0.102 | 0.102 | 0.105 | 0.102 | 0.102 | 0.100 | 0.103 | ||||||||||||
| Specie 15 | 1.233 | 0.115 | 0.115 | 1.560 | 0.100 | 0.090 | 0.102 | 0.102 | 0.105 | 0.102 | 0.102 | 0.100 | 0.103 | 0.000 | |||||||||||
| Specie 16 | 1.233 | 0.115 | 0.115 | 1.560 | 0.100 | 0.090 | 0.105 | 0.105 | 0.105 | 0.102 | 0.102 | 0.100 | 0.106 | 0.002 | 0.002 | ||||||||||
| Specie 17 | 0.113 | 1.253 | 1.253 | 0.074 | 1.240 | 1.167 | 1.138 | 1.138 | 1.128 | 1.237 | 1.237 | 1.234 | 1.288 | 1.133 | 1.133 | 1.133 | |||||||||
| Specie 18 | 0.112 | 1.239 | 1.239 | 0.068 | 1.225 | 1.153 | 1.125 | 1.125 | 1.115 | 1.222 | 1.222 | 1.220 | 1.273 | 1.119 | 1.119 | 1.119 | 0.008 | ||||||||
| Specie 19 | 0.112 | 1.239 | 1.239 | 0.068 | 1.225 | 1.153 | 1.125 | 1.125 | 1.115 | 1.222 | 1.222 | 1.220 | 1.273 | 1.119 | 1.119 | 1.119 | 0.008 | 0.000 | |||||||
| Specie 20 | 1.315 | 0.142 | 0.142 | 1.570 | 0.127 | 0.125 | 0.117 | 0.117 | 0.120 | 0.120 | 0.120 | 0.118 | 0.105 | 0.103 | 0.103 | 0.105 | 1.187 | 1.173 | 1.173 | ||||||
| Specie 21 | 1.310 | 0.144 | 0.144 | 1.576 | 0.129 | 0.125 | 0.117 | 0.117 | 0.121 | 0.122 | 0.122 | 0.120 | 0.108 | 0.105 | 0.105 | 0.108 | 1.187 | 1.173 | 1.173 | 0.002 | |||||
| Specie 22 | 0.116 | 1.733 | 1.733 | 0.095 | 1.309 | 1.633 | 1.359 | 1.359 | 1.342 | 1.581 | 1.581 | 1.579 | 1.388 | 1.603 | 1.603 | 1.603 | 0.112 | 0.106 | 0.106 | 1.592 | 1.597 | ||||
| Specie 23 | 1.311 | 0.147 | 0.147 | 1.711 | 0.127 | 0.125 | 0.113 | 0.113 | 0.115 | 0.119 | 0.119 | 0.117 | 0.105 | 0.107 | 0.107 | 0.109 | 1.187 | 1.173 | 1.173 | 0.000 | 0.002 | 1.751 | |||
| Specie 24 | 0.113 | 1.575 | 1.575 | 0.108 | 1.325 | 1.531 | 1.398 | 1.398 | 1.386 | 1.519 | 1.519 | 1.517 | 1.372 | 1.481 | 1.481 | 1.481 | 0.108 | 0.104 | 0.104 | 1.453 | 1.445 | 0.012 | 1.615 | ||
| Specie 25 | 0.113 | 1.575 | 1.575 | 0.108 | 1.325 | 1.531 | 1.398 | 1.398 | 1.386 | 1.519 | 1.519 | 1.517 | 1.372 | 1.481 | 1.481 | 1.481 | 0.108 | 0.104 | 0.104 | 1.453 | 1.445 | 0.012 | 1.615 | 0.000 | |
| Specie 26 | 0.113 | 1.575 | 1.575 | 0.108 | 1.325 | 1.531 | 1.353 | 1.353 | 1.341 | 1.519 | 1.519 | 1.517 | 1.372 | 1.481 | 1.481 | 1.481 | 0.108 | 0.104 | 0.104 | 1.453 | 1.445 | 0.012 | 1.615 | 0.000 | 0.000 |
 genes in studied bee species (0.2 nucleotide substitution rate per position).")
Phylogenetic tree of nucleotide sequences of cytochrome oxidase I (COI) genes in studied bee species (0.2 nucleotide substitution rate per position).
4.2. Nucleotide Substitutions
a This table shows the percentage of substitution from a row to a column.
b The numbers outside the diagonal represent crossover substitutions.
c The numbers on the diagonal represent transition substitutions.
4.3. Replacement Rate
| Parameters | Numerical Value | Standard Deviation |
|---|---|---|
| dN | 0.030950 | 0.008807 |
| dS | 0.386167 | 0.041641 |
| dN/dS | 0.080146 | 0.000154 |
Abbreviations: dN, nucleotide changes that cause amino acid changes; dS, nucleotide changes that do not affect the resulting amino acid; of dN/dS the process of natural selection.
4.4. Genetic Diversity Indices
| Indices | Theta-W | S | H | Hd | Pi | Eta | K |
|---|---|---|---|---|---|---|---|
| Values | 0.159 | 195 | 4 | 0.966 | 0.259 | 20 | 83.19 |
Abbreviations: Theta-W, mutation rate calculated from all polymorphic sites; S, number of polymorphic loci; H, number of haplotypes; Hd, haplotype frequency; Pi, nucleotide diversity; Eta, total number of mutations; K, nucleotide difference between populations or species (nucleotide divergence).
| Indices | D Tajima Test | CT | MWL | C |
|---|---|---|---|---|
| Values | 1.174 | 0.49 | 80 | 0.396 |
Abbreviations: CT, protection threshold limit; MWL, minimum protection length; C, sequence protection.