1. Context
1.1. Turmeric or Curcuma longa
1.1.1. Health Benefits
1.2. Curcumin

Chemical Structure of Curcumin
1.2.1. Anti-Inflammatory and Antioxidant Effects of Curcumin

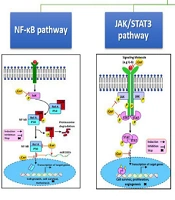
Molecular Targets of Curcumin
1.3. Hyperglycemia and Hyperinsulinemia
1.3.1. Insulin Function and Insulin Resistance
| Substance Name | Role in Insulin Signaling |
|---|---|
| GLUT | Translocated to plasma membrane for glucose influx |
| IL-6 | Impairs/inhibits insulin signaling; suppresses adipogenesis and secretion of adiponectin |
| NF-κB | Influences gene transcription; inhibited by adiponectin |
| PPAR-γ | Stimulates free fatty acid (FFA) catabolism; thiazolidinediones (TZDs) are PPAR-γ agonists |
| TNFα | Main factor for stimulating the secretion of FFAs from adipose tissue into circulation; impairs insulin signaling |
| MCP-1 | Impairs insulin-stimulated glucose uptake; promotes atherosclerosis |
| FFAs | Adipocytes decrease glucose uptake in peripheral tissues by releasing free fatty acids |
| Akt | A key regulator of insulin action and glucose uptake in mammals |
| IRS-1 | Plays a key role in transmitting signals from insulin receptor to intracellular PI3K/Akt pathways and Ras/mitogen activated protein kinase, which eventually mediate various actions of insulin |
| PTP1B | A negative regulator of insulin signaling which targets tyrosine-phosphorylated insulin receptor β and IRS-1 |
| mTOR | Inhibits IRS-1 tyrosine phosphorylation |
aGLUT, Glucose transporter; IL-6, Interleukin-6; NF-κB, Nuclear transcription factor κB; PPAR-γ, Peroxisome proliferator-activated receptor-gamma; TNFα, Tumor necrosis factor alpha; MCP-1,Monocyte chemoattractant protein-1; FFAs, free fatty acids; Akt, protein kinase B; IRS-1, insulin receptor substrate-1; PTP1B, protein–tyrosine phosphatase 1B; mTOR, mammalian target of rapamycin.
1.3.2. Hyperglycemia
1.3.3. Hyperglycemia-Induced Tissue Damage
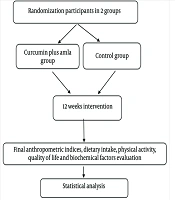
2. Evidence Acquisition
3. Results
3.1. Studies Conducted on the Effects of Curcumin on Blood Glucose Levels and Insulin Resistance
3.1.1. In Vivo/in Vitro Experiments
| In Vivo/in Vitro experiment | Dose, Duration | Results In the End of the Study in Treatment Group | Overall Conclusion | Authors/Publication year | Reference Number |
|---|---|---|---|---|---|
| STZ induced Diabetic Wistar rats (55 mg/kg) | 60 mg/kg Curcumin 14 days | ↑ Gene expression of: acetylcholine esterase, Glut3 Muscarinic M1, M3, α7 nicotinic acetylcholine and insulin receptors | Curcumin plays an important role in regulating the activity of cholinergic and insulin receptors and mechanism of glucose transportation through Glut3, which lead to normalizing the diabetes-mediated cerebellar disorders. Thus, Curcumin has a significant role in a therapeutic application for the prevention or progression of diabetic complications in the cerebellum. | Peeyush,KT and collegues/2009 | (25) |
| high-fat diet+30 mg/kg STZ induced Diabetic rats (in vivo study) | 3 groups 50, 150, or 250mg/kg Curcumin. 7 wk | plasma lipids and glucose ↓,glucose uptake↑, phosphorylated acetyl COA carboxylase in L6 myotubes ↑, phosphorylated AMPK ↑, carnitine palmitoyl transferase 1 ↑, pyruvate dehydrogenase 4 ↓, phosphorylated glycogen synthase (GS) ↓, glycogen synthesis in skeletal muscle ↑, in both in vivo and in vitro studies. | Curcumin enhances muscular insulin resistance by promoting oxidation of fatty acid and glucose, which is, at least in part, mediated through LKB1-AMPK pathway. | Na,LX and collegues/2011 | (26) |
| L6 myotubes (in vitro experiment) | Curcumin at 5, 10, 20, or 40 mol/L | ||||
| high fat diet induced Diabetic Sprague Dawley rats | Curcumin (80 mg/kg) 15 days | hyperglycemia ↓, Improve glucose tolerance, Improve lipid profile, Fasting insulin ↑, Insulin sensivity ↑ | Curcumin exhibited an anti-hyperglycemic effect and improved insulin sensitivity | El-Moselhy,MA and collegues/2011 | (27) |
| Alloxan induced diabetic rats | Turmeric 1 g/kg and Curcumin 0.08 g/kg 21 days | blood sugar ↓, Hb and glycosylated hemoglobin levels ↓ , diabetic oxidative stress ↓ | Curcumin was more effective in attenuating diabetes mellitus related changes than turmeric | Arun,N and colleagues /2002 | (28) |
| Wild-type and ob/ob C57BL/6J mice | Dietary Curcumin (3%) 10 wks. | HbA1c levels ↓, improves glucose tolerance blood glucose levels ↓, adipose, hepatic, and systemic inflammation ↓, Serum adiponectin levels ↑, serum levels of liver and adipocytes MCP-1 ↓ | Curcumin in high oral doses safely treat diabetes in several mouse models of obesity-associated diabetes. Curcumin also greatly improved inflammation at the cellular and biochemical level in white adipose tissue of obese mice. | Weisberg,SP and colleagues /2008 | (29) |
| C57BL/ KsJ-ob/obmice | Curcumin (0.02%, wt/wt) 6 wks | blood glucose ↓, HbA1c levels ↓, improved HOMA-IR and glucose tolerance, plasma insulin ↑, plasma free fatty acid, cholesterol, and triglyceride concentrations hepatic glycogen ↓, skeletal muscle lipoprotein lipase ↑, lipid peroxidation. ↓ | It seems that Curcumin is a potential glucose-lowering agent and antioxidant in type 2 diabetic ob/obmice, but had no effect in non-diabetic db/+ mice. | Seo,KI and colleagues /2008 | (30) |
| type 2 diabetic KK-Ay mice | Turmeric Et-OH Extract 0.2 or 1.0 g/100 g diet 4 Weeks | blood glucose level ↓ | Turmeric Et-OH extract has PPAR-γ ligand-binding activity which probably leads to Adipocyte that finally result in the suppression of an increase in blood glucose levels in type 2 diabetic KK-Ay mice | Kuroda,M and colleagues /2005 | (31) |
| In Vitro Experiments | human pre-adipocytes were cultured with the Turmeric Et-OH Extract 2.0, 5.0, 10, and 20 g/ml | Stimulated adipocyte differentiation | |||
| STZ induced Diabetic rats | 60 mg/kg oral Curcumin 28 days | The expression of both TNF-α and TNF-α receptor 1 ↓, Hyperglycemia ↓ , body weight improved | Curcumin seems to relieve diabetic hyperalgesia, probably through an inhibitory effect on TNF-α and TNF-α receptor 1. | Li,Y and colleagues /2013 | (32) |
| type 2 diabetic KK-Ay mice | turmeric extracts:, 0.1 or 0.5 g of hexane extraction/100 g of diet, 0.1 or 0.5 g of ethanol, extraction from hexane-extraction residue/100 g of diet 0.2 or 1.0 g of ethanol extraction/100 g of diet 4 weeks | E-ext stimulated human adipocyte differentiation turmeric extracts and Curcumin had human peroxisome proliferator-activated receptor-γ (PPAR-γ) ligand-binding activity | Both Curcuminoids and sesquiterpenoids in turmeric exhibit hypoglycemic effects via PPAR-γ activation as one of the mechanisms, and suggest that E-ext including Curcuminoids and sesquiterpenoids has the additive or synergistic effects of both components. | Nishiyama,T and colleagues /2005 | (33) |
| Cell culture of Pancreas and muscle tissues of adult mice | aqueous extract of turmeric (100 g of ground turmeric in 1 L of water) various dose | insulin secretion from mouse pancreatic tissues ↑, glucose uptake ↑ | Water soluble constituents of turmeric exhibit insulin releasing and stimulating actions within in vitro tissue culture conditions | Mohankumar,S and colleagues /2010 | (34) |
| Skeletal muscle isolated from Wistar rats. | 0.1 -1 m Curcumin on cultured C2C12 cells | glucose uptake into skeletal muscle isolated ↑, the membrane protein level of GLUT4 ↑ | Curcumin can activate muscarinic M-1 cholinoceptor (M1-mAChR) at concentrations lower than to scavenge free radicals for increase of glucose uptake into skeletal muscle through PLC-PI3-kinase pathway | Cheng,TC and colleagues /2009 | (35) |
| In vivo: STZ induced Diabetic Sprague–Dawley rats | synthetic analogues of Curcumin )C66) (0.2, 1.0 or 5.0 mg·kg-1), 6 wks. | TNF-a levels and renal inflammatory gene expression ↓ improved histological abnormalities and fibrosis of diabetic kidney | This novel compound is a potential anti-inflammatory agent and might be beneficial for the prevention of diabetic nephropathy; but did not affect the hyperglycemia in these diabetic rats. | Pan,Y and colleagues /2012 | (36) |
| In vitro :Primary peritoneal macrophages (MPM), prepared from C57BL/6 mice | Treated with HG (high glucose) in the presence or absence of C66. | Inhibited HG-stimulated production of TNF-α , NO, IL-1β, IL-6, IL-12, COX-2 and iNOS mRNA transcription, and the activation of JNK/NF-κB signalling | |||
| Male Golden-Syrian hamsters | Curcumin (0.05-g/100-g diet) on a high-fat diet 10 weeks. | levels of free fatty acid, total cholesterol, triglyceride ↓, leptin ↓, HOMA-IR ↓, HDL-C ↑, apolipoprotein (apo) A-I ↑, paraoxonase activity in plasma ↑, fatty acid β-oxidation activity ↑, fatty acid synthase ↓, 3-hydroxy-3-methylglutaryl coenzyme A reductase ↓, acyl coenzyme A:cholesterol acyltransferase ↓, lipid peroxide levels in the erythrocyte and liver ↓ | Curcumin showes an obvious hypolipidemic effect by increasing plasma paraoxonase activity, ratios of high-density lipoprotein cholesterol to total cholesterol and of apo A-I to apo B, and hepatic fatty acid oxidation activity with simultaneous inhibition of hepatic fatty acid and cholesterol biosynthesis in high-fat–fed hamsters. | Jang,EM and colleagues /2008 | (37) |
| low dose of STZ + high energy intake induced diabetic wistar rats | Curcumin 100 or 200 mg/kg/ 16 weeks | myocardial dysfunction ↓, cardiac fibrosis ↓ , AGEs accumulation ↓ ,oxidative stress ↓,inflammation ↓,apoptosis ↓, phosphorylation of Akt and GSK-3b ↑ Activation Akt/GSK-3b Signaling Pathway ↑ blood glucose levels ↓, TG ↓ | Curcumin may have great therapeutic effect in the treatment of DCM, and perhaps other cardiovascular disorders, by enhancing fibrosis, oxidative stress, inflammation and cell death | Yu,W and colleagues /2012 | (38) |
| STZ (65 mg/kg) induced Diabetic rats | Curcumin (15 and 30 mg/kg) 2 weeks | renal dysfunction ↓, oxidative stress ↓ | Perhaps anti-oxidative mechanism being responsible for the nephroprotective effect of Curcumin. | Sharma,S and colleagues /2006 | (39) |
| C57BL6 mice (5-6 weeks old) | 24 weeks either a high-fat diet (45% fat) or a low-fat diet (10% fat) together with CNB-001 (40 mg/kg/d) | a novel neuroprotective curcuminoid CNB-001: body weight gain ↓, serum triglycerides ↓, IL-6 ↓, Improved insulin signaling: p-Akt (phosphoprotein kinase B) ↑, p-IR (phosphoinsulin receptor) β ↑, endoplasmic reticulum (ER) stress ↓, proteintyrosine phosphatase1B (PTP1B) ↓ | CNB-001 alleviates obesity-induced glucose intolerance and represents a potential candidate for further development as an anti-diabetic agent | Panzhinskiy E and colleagues /2014 | (16) |
a STZ, Streptozotocin; HG, high glucose; p-Akt, phosphoprotein kinase B ; p-IR, phosphoinsulin receptor β; ER, endoplasmic reticulum; PTP1B, protein–tyrosine phosphatase1B; TG,Triglyceride.
3.1.2. Curcumin, Cell Function and Insulin
3.1.3. The Curcumin Reduction Effects on Hyperglycemia-Induced Circulating ICAM-1, VCAM-1
3.2. Human Clinical Trials of Curcumin

Antihyperglycemic and Anti-Insulin Resistance Effects of Curcumin.
| patients | Dose, Duration | Results In the End of the Study in Treatment Group | Overall Conclusion | Authors/Publication year | Reference Number |
|---|---|---|---|---|---|
| 100 Overweight/obese type 2 diabetic patients | Curcuminoids (300 mg/day) 3 months | fasting blood glucose ↓, insulin resistance index (HOMA-IR) ↓, serum total FFAs ↓, serum triglycerides ↓, LPL activity ↑ | These findings indicate a glucose-lowering effect of Curcuminoids in type 2 diabetes, which is partially due to decrease in serum FFAs, which may result from promoting fatty acid oxidation and utilization | Na LX and colleagues/ 2012 | (46) |
| subjects (n = 240) with criteria of prediabetes | six Curcumin capsules per day (Each capsule has Curcuminoid content of 250mg) 9months | 16.4% of subjects in the placebo group were diagnosed with T2DM, whereas none were diagnosed with T2DM in the Curcumin-treated group the Curcumin-treated group showed: a better overall function of βcells HOMA-β ↑, C-peptide ↓, HOMA-IR ↓, Adiponectin ↑ | Curcumin intervention in a pre-diabetic population significantly lowered the number of pre diabetic individuals who eventually developed T2DM. In addition, the Curcumin treatment appeared to improve overall function of β-cells, with very minor adverse effects. | Chuengsamarn Sand colleagues /2012 | (47) |
| Eleven healthy subjects, aged 21–38 y with normal fasting blood glucose (100 mg/dL) and total cholesterol (200 mg/dL) concentrations | a randomly assigned, crossover design: 6 subjects 3 .0g cinnamon 5subjects 2.8 g turmeric per day each treatment 4-wk | no significant changes in fasting plasma glucose or lipids in conjunction with the 4-wk periods of each treatment. | Maybe turmeric supplementation in healthy nondiabetic subjects did not reflect blood glucose-lowering effect | Tang M and colleagues/ 2008 | (48) |
| 60 subjects, 20 years old and above, who were diagnosed mild to moderate elevated ALT levels with normal glucose levels(91.9 ± 9.7) | fermented turmeric powder 3.0 g per day 12 weeks | reduction in ALT,AST levels no significant changes in blood glucose level | FTP is effective and safe, generally well-tolerated without severe Adverse events, in the treatment of subjects with elevated ALT levels over a 12 weeks period | Kim SW and colleagues /2013 | (49) |
| Fourteen healthy subjects in a crossover trial. | 75 g oral glucose tolerance test (OGTT) together with capsules containing a placebo or C. longa (6 g) | no significant effect on the glucose response The insulin AUCs were significantly higher | The ingestion of 6 g C. longa increased postprandial serum insulin levels, but did not seem to affect plasma glucose levels or GI, in healthy subjects. The results indicate that C. longa may have an effect on insulin secretion | Wickenberg J and colleagues/ 2010 | (47) |
a HOMA-IR, insulin resistance index; OGTT,glucose tolerance test.