



1. Background
2. Objectives
3. Methods
3.1. Cells
3.2. Virus Strain
3.3. Primary Screening for Antiviral Activities
3.3.1. Cytotoxicity Assay
3.3.2. Virus Yield Reduction Assay
3.4. Evaluation of the Mechanism of Action
3.4.1. Virucidal Assay
3.4.2. Adsorption Assay
3.4.3. Penetration Assay
3.4.4. Time of Addition Assay
3.5. Real-Time PCR
3.6. Quantitative Reverse-Transcriptase PCR
| Gene | Primer Name | Sequence (5’→3’) | |
|---|---|---|---|
| UL27 | gB | Forward | AAACCGAAAAACCCACCGCC |
| Reverse | TGTTCTCCGCCTTGATGTCC | ||
| US6 | gD | Forward | GCCCCGCTGGAACTACTATG |
| Reverse | TTATCTTCACGAGCCGCAGG | ||
| UL22 | gH | Forward | GGTTTATGGTTCGTGGGGGT |
| Reverse | CTGTCTGCTCAGTCCAGTCG | ||
| UL36 | VP1/2 | Forward | CGGGTCAAAAAGGTATGCGG |
| Reverse | TGTCGTACACGCTCCTAACC | ||
| UL48 | VP16 | Forward | TTTGACCCGCGAGATCCTAT |
| Reverse | GCTCCGTTGACGAACATGAA | ||
| α4 | ICP4 | Forward | CGACACGGATCCACGACCC |
| Reverse | GATCCCCCTCCCGCGCTTCGTCCG | ||
| UL29 | ICP8 | Forward | CGACAGTAACGCCAGAAG |
| Reverse | GGAGACAAAGCCCAAGAC | ||
| UL54 | ICP27 | Forward | ATGTGCATCCACCACAACCT |
| Reverse | TCCTTAATGTCCGCCAGACG | ||
| US1 | ICP22 | Forward | CGCCGCAGAAGACCGCAAGT |
| Reverse | TGTCGCTGCACGGATAGGG | ||
| GAPDH | GAPDH | Forward | GGTGGTCTCCTCTGACTTCAACA |
| Reverse | GTTGCTGTAGCCAAATTCGTTGT | ||
Abbreviations: GAPDH, glyceraldehyde-3-phosphate dehydrogenase; US, unique short ;UL, unique long; ICP, infected cell protein; VP, viral protein; g, glycoprotein.
3.7. SDS-PAGE and Immunoblotting Assays
3.8. Statistics
4. Results
4.1. Identification of 10-HCPT as Inhibitors of HSV Infection
-10-hydroxycamptothecin as inhibitors of herpes simplex virus type 1 infection. Primary screening of 133 compounds from the natural product library in a single dose of 5 μM. Inhibitory effects were calculated as percentage cytopathogenic effect inhibition. Hits were selected with a cutoff of 60% cytopathogenic effect inhibition (dotted line).")
Identification of triptolide and (S)-10-hydroxycamptothecin as inhibitors of herpes simplex virus type 1 infection. Primary screening of 133 compounds from the natural product library in a single dose of 5 μM. Inhibitory effects were calculated as percentage cytopathogenic effect inhibition. Hits were selected with a cutoff of 60% cytopathogenic effect inhibition (dotted line).
4.2. Cytotoxicity of 10-HCPT
Abbreviations: CC50, 50% cytotoxic concentration; EC50, half maximal effective concentration.
a Values obtained by the plaque reduction assay.
b Values obtained by WST-1 on A549 cells.
c Values obtained by WST-1 on primary cells.
4.3. Plaque Reduction Assays and Determination of the EC50
4.4. Evaluation of the Possible Antiviral Mechanism
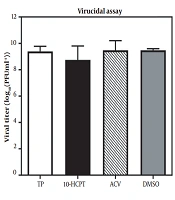
-10-hydroxycamptothecin-mediated inhibition of herpes simplex virus type 1. The virucidal efficacy of (S)-10-hydroxycamptothecin. Herpes simplex virus type 1 was mixed with acyclovir, dimethyl sulfoxide (0.10%; consisting of dimethyl sulfoxide, virus, and cell), and (S)-10-hydroxycamptothecin (6 × EC<sub>50</sub>) and then incubated at 25°C (room temperature) for 1 hour. The virus titers were indicated by a plaque reduction assay. The data were obtained by repeated experiments. Error bars demonstrate SDs from 3 times independent experiments.")
The mechanism study of (S)-10-hydroxycamptothecin-mediated inhibition of herpes simplex virus type 1. The virucidal efficacy of (S)-10-hydroxycamptothecin. Herpes simplex virus type 1 was mixed with acyclovir, dimethyl sulfoxide (0.10%; consisting of dimethyl sulfoxide, virus, and cell), and (S)-10-hydroxycamptothecin (6 × EC50) and then incubated at 25°C (room temperature) for 1 hour. The virus titers were indicated by a plaque reduction assay. The data were obtained by repeated experiments. Error bars demonstrate SDs from 3 times independent experiments.
-10-hydroxycamptothecin on the virus, A, adsorption; and B, penetration steps of entry. This procedure was described in the Methods Section. The data represent the results of independent experiments performed 3 times. Statistically, the significant value between the treated group and dimethyl sulfoxide-treated group as control was evaluated by a Student <i>t</i> test as follows: ns, ***, P < 0.001 or ****, P < 0.0001.")
The effect of (S)-10-hydroxycamptothecin on the virus, A, adsorption; and B, penetration steps of entry. This procedure was described in the Methods Section. The data represent the results of independent experiments performed 3 times. Statistically, the significant value between the treated group and dimethyl sulfoxide-treated group as control was evaluated by a Student t test as follows: ns, ***, P < 0.001 or ****, P < 0.0001.
, and (S)-10-hydroxycamptothecin (3 × EC<sub>50</sub>) were added to the cells at the demonstrated study time points of infection (before and after infection). The infected cells were lysed by doing 3 steps of freezing and thawing. The treated viruses in specific time courses were further evaluated by titration on the Vero cell. The data were assessed by virus yield reduction of cell lysates and represent the mean of a 3-independent experiment (± SE). (∗∗∗) demonstrated significant statistical differences between the treated groups (P < 0.0001). Analysis of variance/Dunnett tests were carried out as appropriate.")
Time-of-addition study. The A549 cells infected with herpes simplex virus type 1 at MOI of 0.1, acyclovir (0.01 μg/mL), and (S)-10-hydroxycamptothecin (3 × EC50) were added to the cells at the demonstrated study time points of infection (before and after infection). The infected cells were lysed by doing 3 steps of freezing and thawing. The treated viruses in specific time courses were further evaluated by titration on the Vero cell. The data were assessed by virus yield reduction of cell lysates and represent the mean of a 3-independent experiment (± SE). (∗∗∗) demonstrated significant statistical differences between the treated groups (P < 0.0001). Analysis of variance/Dunnett tests were carried out as appropriate.
4.5. Yields of Viral Nucleic Acid in the Presence of 10-HCPT
-10-hydroxycamptothecin (a), in the presence or absence of (S)-10-hydroxycamptothecin, compared to the dimethyl sulfoxide control (consisting of the cells, dimethyl sulfoxide, and virus). Data are presented as mean values of triplicate with its SD. Significant differences were found among herpes simplex virus type 1 DNA levels up to 8 hours after infection from (S)-10-hydroxycamptothecin concerning dimethyl sulfoxide controls (consisting of the cells, dimethyl sulfoxide, and virus). (∗∗∗) indicates significant statistical differences between the treated groups and the dimethyl sulfoxide control (P < 0.0001). Analysis of variance/Dunnett tests were carried out as appropriate.")
The real-time polymerase chain reaction was performed to evaluate the number of herpes simplex virus type 1 DNA copies after treating with (S)-10-hydroxycamptothecin (a), in the presence or absence of (S)-10-hydroxycamptothecin, compared to the dimethyl sulfoxide control (consisting of the cells, dimethyl sulfoxide, and virus). Data are presented as mean values of triplicate with its SD. Significant differences were found among herpes simplex virus type 1 DNA levels up to 8 hours after infection from (S)-10-hydroxycamptothecin concerning dimethyl sulfoxide controls (consisting of the cells, dimethyl sulfoxide, and virus). (∗∗∗) indicates significant statistical differences between the treated groups and the dimethyl sulfoxide control (P < 0.0001). Analysis of variance/Dunnett tests were carried out as appropriate.
4.6. 10-HCPT Effect on the Expression of HSV-1 IE, E, and L late Genes
-10-hydroxycamptothecin (3 × EC<sub>50</sub>) on the expression of herpes simplex virus type 1 genes (immediate-early, and late). A, A549 cells were infected with herpes simplex virus type 1 at 0.1 MOI and then treated with or without 3 × EC<sub>50</sub> of each compound. RNA was extracted at specified intervals, and a quantitative real-time polymerase chain reaction was performed using messenger RNA-specific primers. The gene transcription level at each time point was initially normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and then compared with the virus control group into 24 hours after infection.")
Effects of (S)-10-hydroxycamptothecin (3 × EC50) on the expression of herpes simplex virus type 1 genes (immediate-early, and late). A, A549 cells were infected with herpes simplex virus type 1 at 0.1 MOI and then treated with or without 3 × EC50 of each compound. RNA was extracted at specified intervals, and a quantitative real-time polymerase chain reaction was performed using messenger RNA-specific primers. The gene transcription level at each time point was initially normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and then compared with the virus control group into 24 hours after infection.