




1. Background
As an opportunistic pathogen, Mycobacterium kansasii is a well-known member of slow-growing nontuberculous mycobacteria (NTM) (1). Mycobacterium kansasii was first introduced by Buhler and Pollack in the 1950s (2). Among NTM species, M. kansasii is the most virulent mycobacterium after the M. avium complex and causes pulmonary and extrapulmonary infections (3). Reports from various regions of Europe, Asia, Brazil, and the USA indicate the prevalence of this pathogen (4, 5). To date, among the seven subtypes of M. kansasii, only subtypes I and II are accepted as clinically pathogenic. The diseases caused by M. kansasii and M. tuberculosis share common symptoms, including cough, chest pain, and dyspnea (6, 7).
In cases of verified highly susceptible M. kansasii infections, the American Thoracic Society/Infectious Diseases Society of America (ATS/IDSA) recommends the use of isoniazid (INH), rifampin (RIF), and ethambutol (EMB) for at least one year (8, 9). In instances of resistance to the preferred first-line antibiotics, clarithromycin (CLA), amikacin (AMK), rifabutin (RFB), linezolid (LZD), and fluoroquinolones (FQs) can be used as second-line and alternative drugs. Rifampin and CLR drug susceptibility testing (DST) is the first step of susceptibility tests for M. kansasii recommended by the Clinical and Laboratory Standards Institute (CLSI).
In cases of resistance to RIF, the guideline recommends determining susceptibility to first- and second-line drugs such as EMB, RFB, ciprofloxacin (CIP), streptomycin (STR), AMK, trimethoprim/sulfamethoxazole (TMP-SMZ), moxifloxacin (MXF), and LZD (10, 11). Identifying the genes that cause drug resistance in M. kansasii and understanding the resistance mechanisms are important for administering correct treatment. Mutations in the genes rpoB, rbpA, gyrA, gyrB, and rrl in M. kansasii could cause resistance to RIF, CIP, and CLR drugs, respectively (12). A review of the literature indicates that studies investigating drug resistance in M. kansasii both phenotypically and genotypically are rare. In the case of Turkey, the current research is the first comprehensive study of drug resistance in M. kansasii isolates.
2. Objectives
The present study aimed to evaluate the drug susceptibility of 40 M. kansasii isolates from Turkey. Additionally, we investigated mutations in resistant M. kansasii isolates associated with drug resistance.
3. Methods
3.1. Mycobacterial Isolates
In the initial phase of the study, 48 M. kansasii isolates were examined, obtained from respiratory tract specimens [such as sputum, bronchoalveolar lavage (BLA), and bronchial aspirates] collected between 2021 and 2023 from the Tuberculosis Reference Laboratory of the General Directorate of Public Health in Ankara, Turkey. These isolates were cultured on Lowenstein-Jensen (LJ) medium and incubated for 6 weeks at 37°C. Forty M. kansasii isolates (13), confirmed through both routine culture and molecular tests, were selected, and isolates that did not grow on the culture medium were excluded.
3.2. Antimicrobial Susceptibility Testing
Antimycobacterial susceptibility testing was performed and interpreted according to CLSI M24-A2 guidelines (14) and the manufacturer’s instructions. Sensititre® SLOMYCO commercial plates (TREK Diagnostic Systems, West Sussex, UK) including 13 drugs — AMK (1 - 64 mg/mL); CIP (0.12 - 16); doxycycline (DOX) (0.12 - 16); ethionamide (ETH) (0.3 - 20); LZD (1 - 64); EMB (0.5 - 16 mg/mL); STR (0.5 - 64 mg/mL); MXF (0.12 - 8 mg/mL); RIF (0.12 - 8); INH (0.25 - 8 mg/mL); RFB (0.25 - 8 mg/mL); CLA (0.06 - 64 mg/mL); TMP-SMZ (0.12 - 8 mg/mL) — were used (15). The Sensititre SLOMYCO plates were incubated at 37°C until the growth control showed sufficient growth after 7 - 14 days of incubation. Readings for M. kansasii were taken visually using an inverted mirror, and wells with no growth were accepted as minimum inhibitory concentrations (MICs) (16). Due to the constraints of the SLOMYCO panel system used in this study, which is optimized for quality control with a single reference strain, additional resistant control strains could not be included. Mycobacterium kansasii ATCC 12478 was used as a reference strain for MIC testing.
3.3. DNA Extraction
DNA was extracted from the isolates using the FluoroLyse kit (Hain Lifescience, Nehren, Germany) according to the manufacturer’s instructions. In the first stage, 300 μL of FluoroLyse was placed in a microcentrifuge tube. Following the transfer of 2 - 3 colonies from Lowenstein-Jensen medium, the tube was boiled at 95°C for 5 minutes. Then, after a short spin, 100 μL of neutralization buffer (F-NB) was added to the tube, and the mixture was vortexed for 5 seconds and centrifuged at 13,000 g for 5 minutes. Finally, the remaining supernatant was transferred to microcentrifuge tubes.
3.4. Polymerase Chain Reaction
Polymerase chain reaction (PCR) was used to amplify the target genes CLR, CIP, and RIF in M. kansasii strains using primers as described in Table 1. Primers used for amplification of the rpoB and rbpA genes were designed by the authors based on M. kansasii sequences available in GenBank and optimized in-house for this study, whereas primers targeting gyrA, gyrB, and rrl genes were adopted from previously published studies (3). For the amplification of rrl, gyrA, and gyrB, a 30 μL reaction mixture was prepared, consisting of 15 µL of 2X Taq Premix (Genet Bio Inc.), 1.5 µL each of the 10 pmol forward and reverse primers, and 12 µL of genomic DNA. The amplification of rrl, gyrA, and gyrB genes was performed using the Labcycler Basic (SensoQuest GmbH) thermocycler under the following conditions: An initial denaturation at 95°C for 5 minutes, followed by 35 cycles of denaturation at 95°C for 30 seconds, annealing at a temperature range of 58°C to 60°C for 30 seconds, and extension at 72°C for 90 seconds. A final extension was carried out at 72°C for 5 minutes.
For the amplification of rpoB and rbpA, a 20 μL reaction mixture was prepared, consisting of 10 µL of 2X Phanta Max Master Mix (Vazyme), 1 µL each of the 10 pmol forward and reverse primers, and 8 µL of genomic DNA. The amplification of these genes was performed using the Labcycler Basic (SensoQuest GmbH) thermocycler with the following protocol: An initial denaturation at 95°C for 3 minutes, followed by 35 cycles of denaturation at 95°C for 30 seconds, annealing at 46°C for rpoB or 52°C for rbpA for 30 seconds, and extension at 72°C for 45 seconds for rbpA and 4 minutes for rpoB. A final extension was carried out at 72°C for 5 minutes. The PCR products were run on a 1.5% agarose gel electrophoresis (120 V, 60 min) stained with Ultra GelRed Stain (Vazyme, Nanjing, China) and using a 1 kb plus DNA ladder (Thermo Scientific). The DNA bands were observed under UV light in a BioRad ChemiDoc MP Imaging System (Universal Hood III, USA).
| Genes and Primers | Size (bp) | References |
|---|---|---|
| gyrA | 459 | (3) |
| F: ATTCTGCCGAACGGATCGAG | ||
| R: CGACCGCGTTATCCGAATTG | ||
| gyrB | 762 | (3) |
| F: TGGGCAACACCGAGGTGAAG | ||
| R: ACGGGTCCATGGTGGTTTCC | ||
| rrl | 1110 | (3) |
| F: CCAGGTCTGGCCTATCGAAC | ||
| R: CGGGATTCGGTCGCAGAAAC | ||
| rpoB | 3603 | Designed by our research team |
| F: ACGCATCTTGGCTGATTT | ||
| R: TTCATCGAAGAAGTTGACGT | ||
| RbpA | 550 | Designed by our research team |
| F: GGTATCATCGATGGTCGC | ||
| R: TGGGGTATTGAGGGCAGA |
3.5. DNA Sequencing (Sanger Sequencing)
The amplification products were purified using Kapa Pure Beads (Roche Sequencing), and cycle sequencing was performed using the BrilliantDye™ Terminator (v3.1) Cycle Sequencing Kit (NimaGen, Netherlands) according to the manufacturer’s instructions. Cycle sequencing was performed (after a first denaturation step at 96°C for 1 minute) for 30 cycles at 96°C for 10 seconds, at 50°C for 5 seconds, with a final extension at 60°C for 4 minutes and a 4°C final hold. The PCR amplicons, after purification (Omega Bio-tek’s Mag-Bind SeqDTR), were sequenced on the ABI 3500 Genetic Analyzer (Applied Biosystems). Sequence analysis was performed with the Molecular Evolutionary Genetics Analysis (MEGA) software version 11 (www.megasoftware.net). The sequences obtained for 40 M. kansasii isolates were compared to the M. kansasii ATCC 12478 reference strain.
3.6. Oxford Nanopore Technologies Amplicon DNA Sequencing (Next Generation Sequencing)
RpoB and rbpA PCR amplicons of the 36 RIF-resistant M. kansasii samples were quantified using a Qubit Flex Fluorometer with the 1X dsDNA HS kit (Invitrogen, USA). According to the ONT amplicon sequencing library preparation protocol (SQK-NBD114.96), the DNA concentration of each sample was adjusted to 200 fmol (130 ng for 1 kb amplicons). Subsequently, each sample was made up to a final volume of 11.5 µL using nuclease-free water. Also, 3.5 μL of end-prep reagents master mix (1 μL DNA control sample (DCS), 1.75 μL Ultra II End-prep Reaction Buffer, 0.75 μL Ultra II Enzyme Mix) was added to a well of each sample. After incubation, in a new 96-well plate, 1.25 μL Native Barcode (NB01-96), 5 μL Blunt/TA Ligase Master Mix, and 3 μL nuclease-free water were added to 0.75 μL End-prepped DNA.
Twenty minutes after incubation, 1 μL EDTA was added to each well, and then 36 barcoded samples were pooled in a 1.5 mL tube, and purification was carried out using AMPure XP Beads (AXP). For the adapter ligation step reaction mix, in a 0.2 PCR tube, 30 μL of eluate from the previous step, 5 μL of native adapter (NA), 50 μL of 10X ligase buffer, and 5 μL of T4 DNA ligase (Vazyme-C301-01 40,000 U) were mixed by performing pipette mixes. After incubation at 37°C for 10 minutes, a cleaning step was performed using Short Fragment Buffer (SFB) rather than 80% ethanol to wash and AXP beads (Beckman Coulter, USA). According to DNA library fragment size, in a new 1.5 mL tube, 50 fmol from 12 μL of eluate containing the DNA library, 37.5 μL Sequencing Buffer (SB), and 25.5 μL Library Beads (LIB) were mixed and loaded to the R10.4.1 flow cell with 1352 pores. Amplicon sequencing was performed in an ONT sequence device (GridION), and the run was stopped after 20 hours and washed using a Flow Cell Wash Kit (EXP-WSH002) according to the ONT’s protocol.
3.7. ONT Amplicon Sequencing Data Analysis
Following sequencing, the results obtained in POD5 format were converted to FASTQ format using the latest version of the Dorado software, which performed both base-calling and de-multiplexing. Barcode and adapter sequences were removed using Dorado, and low-quality bases were further processed with Trimmomatic (v0.39). The first 15 bases of each read were trimmed due to lower quality; reads with a quality score below 20 were excluded from the analysis. The processed reads were then aligned to the M. kansasii reference genes rpbA and rpoB using the minimap2 tool within the Geneious Prime software (v23.2.2). Variants were identified and reported using the "Find Variations & SNPs" tool in Geneious Prime.
3.8. Statistical Analysis
The analyses were performed using Statistical Package for the Social Sciences (SPSS) version 23. Descriptive statistics for continuous variables were presented as medians with interquartile ranges (IQR), while those for categorical variables were presented as frequencies (no.) and percentages (%). For bivariate analysis, Pearson correlation coefficients (R) were calculated to assess the relationship between the MIC90 values of nine antimicrobial agents. The strength of the correlation was evaluated as follows: 0.00 - 0.19 (very weak); 0.20 - 0.39 (weak); 0.40 - 0.59 (moderate); 0.60 - 0.79 (strong); 0.80 - 1.0 (very strong). A P-value of less than 0.05 was considered statistically significant.
4. Results
The study group consisted of 40 patients, with 92.5% (n = 37) being male and 7.5% (n = 3) being female. The mean age of the patients was 49.5 years, with an IQR of 37.0 to 24.0 years. Most of the samples were from sputum or phlegm, accounting for 82.5% (n = 33). Other sample types included BAL in 12.5% (n = 5) of the cases, and other types in 5% (n = 2) (Table 2). Geographically, the patients were distributed across various regions of Turkey, with the highest numbers from Bursa at 22.5% (n = 9), Adana at 15.0% (n = 6), and Izmir at 15.0% (n = 6). Other locations had fewer patients and were grouped under ‘Other regions’ due to lower case counts (Table 2).
| Characteristics (n = 40) | Values |
|---|---|
| Sex | |
| Female | 3 (7.5) |
| Male | 37 (92.5) |
| Age [y; median (IQR)] | 49.5 (37.0 - 24.0) |
| Sample type | |
| Sputum | 33 (82.5) |
| BAL | 5 (12.5) |
| Other (bronchial aspirate, etc.) | 2 (5.0) |
| Location | |
| Bursa | 9 (22.5) |
| Adana | 6 (15.0) |
| Izmir | 6 (15.0) |
| Other regions | 19 (47.5) |
Abbreviation: BAL, bronchoalveolar lavage.
a Values are expressed as No. (%) unless otherwise indicated.
Table 3 presents the susceptibility test results of M. kansasii to the nine agents. According to the breakpoints of drugs in the CLSI guidelines (M24-A2), all 40 M. kansasii isolates were susceptible to CLR, AMK, RFB, MXF, and LZD. Furthermore, all 40 isolates (100%) were resistant to EMB, 36 isolates (90.0%) were resistant to RIF, and 36 isolates (90.0%) were resistant to CIP. Additionally, 28 isolates (70.0%) were resistant to TMP-SMZ.
| Antimicrobial Agent (n = 9) | MIC90 Values (µg/mL), Median (IQR) | R, No. (%) |
|---|---|---|
| AMK | 2.00 (2.00 - 3.50) | 0 (0.0) |
| CLA | 0.25 (0.25 - 0.50) | 0 (0.0) |
| EMB | 8.00 (8.00 - 16.00) | 40 (100.0) |
| LZD | 4.00 (2.00 - 4.00) | 0 (0.0) |
| MXF | 0.25 (0.25 - 0.25) | 0 (0.0) |
| RFB | 0.50 (0.50 - 1.00) | 0 (0.0) |
| RIF | 3.00 (2.00 - 4.00) | 36 (90.0) |
| CIP | 8.00 (4.00 - 8.00) | 36 (90.0) |
| TMP-SMZ | 8.00/152.00 (0.25/4.75 - 8.00/152.00) | 28 (70.0) |
Abbreviations: MIC, minimum inhibitory concentration; R, resistant; AMK, amikacin; CLA, clarithromycin; LZD, linezolid; MXF, moxifloxacin; RFB, rifabutin; CIP, ciprofloxacin; RIF, rifampin; EMB, ethambutol; TMP-SMZ, trimethoprim/sulfamethoxazole.
Previous research has revealed a relationship between rrl gene mutations and CLA resistance (17). Moreover, RIF resistance and CIP resistance are associated with mutations in the rpoB gene and the gyrA and gyrB genes, respectively. Mutations in the rbpA gene may affect mycobacterial rifampicin sensitivity. We studied M. kansasii resistance mechanisms using Sanger and targeted nanopore sequencing. The study did not find any changes in the rrl, gyrA, or gyrB genes that are usually linked to resistance to CIP or CLA. In this study, we also investigated mutations in the rpoB and rbpA genes using nanopore sequencing technology.
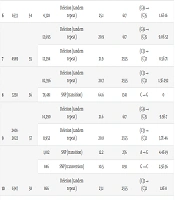
Thirty-six out of 40 M. kansasii isolates (90%) demonstrated resistance to RIF. ONT targeted sequencing was performed on 36 resistant isolates; however, no mutations were identified in five of these isolates, and six others were excluded from analysis due to insufficient sequencing data. ONT sequencing revealed the presence of multiple mutations in 18 isolates and single mutations in another 5. Mutations in the rifampin resistance-determining region (RRDR) were discovered most often at codons 447 and 457. Additionally, ONT sequencing revealed, for the first time in this study, mutations at nucleotides 226, 617, 1191, and 2555 outside the RRDR (Table 4). The M. kansasii ATCC 12478 DNA-directed RNA polymerase beta subunit (rpoB) gene, partial cds (GenBank: KY933079.1), was used as the reference genome for alignment.
| C | Samples | Barcodes | Coverages | Polymorphism Types | Variant Frequency | Nucleotide No. | Change | Variant P-Value |
|---|---|---|---|---|---|---|---|---|
| 1 | 5076 | 49 | 19,482 | Deletion (tandem repeat) | 21.3 | 2555 | (C)3 → (C)2 | 2.5E-90 |
| 2 | 4959 | 50 | 21,828 | Deletion (tandem repeat) | 22.2 | 617 | (G)6 → (G)5 | 2.2E-16 |
| 20,265 | Deletion (tandem repeat) | 21.0 | 2555 | (C)3 → (C)2 | 2.5E-90 | |||
| 3 | 6312 | 51 | 24,166 | Deletion (tandem repeat) | 22.3 | 617 | (G)6 → (G)5 | 2.1E-20 |
| 21,753 | Deletion (tandem repeat) | 21.6 | 2555 | (C)3 → (C)2 | 1.4E-108 | |||
| 19,834 | SNP (transition) | 68 | 1341 | C → G | 0 | |||
| 18,602 | SNP (transition) | 62 | 1371 | G → C | 0 | |||
| 4 | 4538 | 52 | 26,650 | Deletion (tandem repeat) | 21.2 | 2555 | (C)3 → (C)2 | 3.6E-115 |
| 5 | 7027 | 53 | 21,016 | Deletion (tandem repeat) | 22.2 | 617 | (G)6 → (G)5 | 2.5E-16 |
| 6 | 6533 | 54 | 4,328 | Deletion (tandem repeat) | 25.1 | 617 | (G)6 → (G)5 | 1.6E-16 |
| 7 | 4989 | 55 | 12,695 | Deletion (tandem repeat) | 20.9 | 617 | (G)6 → (G)5 | 9.0E-52 |
| 12,394 | Deletion (tandem repeat) | 21.9 | 2555 | (C)3 → (C)2 | 8.5E-71 | |||
| 8 | 3258 | 56 | 82,396 | Deletion (tandem repeat) | 20.7 | 2555 | (C)3 → (C)2 | 1.3E-292 |
| 78,481 | SNP (transition) | 64.6 | 1341 | C → G | 0 | |||
| 9 | 2401-2022 | 57 | 14,290 | Deletion (tandem repeat) | 21.6 | 617 | (G)6 → (G)5 | 9.9E-7 |
| 11,952 | Deletion (tandem repeat) | 20.8 | 2555 | (C)3 → (C)2 | 3.7E-46 | |||
| 10 | 6347 | 58 | 1,012 | SNP (transition) | 12.2 | 276 | A → G | 4.4E-19 |
| 846 | SNP (transversion) | 10.5 | 1191 | G → C | 2.5E-36 | |||
| 866 | Deletion (tandem repeat) | 23.1 | 2555 | (C)3 → (C)2 | 1.8E-8 | |||
| 11 | 3254 | 59 | 22,507 | Deletion (tandem repeat) | 25.5 | 617 | (G)6 → (G)5 | 3.2E-91 |
| 21,230 | Deletion (tandem repeat) | 23.3 | 2555 | (C)3 → (C)2 | 1.9E-32 | |||
| 24,255 | SNP (transition) | 49.1 | 1341 | C → G | 0 | |||
| 12 | 4720 | 60 | 10,048 | Deletion (tandem repeat) | 26.8 | 617 | (G)6 → (G)5 | 1.0E-61 |
| 9,406 | Deletion (tandem repeat) | 23.7 | 2555 | (C)3 → (C)2 | 3.3E-19 | |||
| 13 | 8951 | 61 | 17,347 | Deletion (tandem repeat) | 26.7 | 226 | (G)6 → (G)5 | 5.8E-102 |
| 13,929 | Deletion (tandem repeat) | 26.0 | 617 | (G)6 → (G)5 | 2.9E-67 | |||
| 13,003 | Deletion (tandem repeat) | 23.9 | 2555 | (C)3 → (C)2 | 5.6E-29 | |||
| 14 | 5640 | 62 | 17,652 | Deletion (tandem repeat) | 25.1 | 617 | (G)6 → (G)5 | 2.0E-61 |
| 16,425 | Deletion (tandem repeat) | 23.9 | 2555 | (C)3 → (C)2 | 4.5E-36 | |||
| 15 | 3700 | 63 | 29,532 | Deletion (tandem repeat) | 26.2 | 617 | (G)6 → (G)5 | 2.6E-148 |
| 27,717 | Deletion (tandem repeat) | 23.7 | 2555 | (C)3 → (C)2 | 8.6E-52 | |||
| 24,156 | SNP (transition) | - | 1341 | C → G | 0 | |||
| 20,603 | SNP (transition) | - | 1371 | G → C | 0 | |||
| 16 | 6529 | 64 | 4,783 | Deletion (tandem repeat) | 25.2 | 2555 | (C)3 → (C)2 | 1.0E-18 |
| 17 | 3650 | 65 | 42,270 | Deletion (tandem repeat) | 25.4 | 617 | (G)6 → (G)5 | 2.2E-165 |
| 39,300 | Deletion (tandem repeat) | 24.1 | 2555 | (C)3 → (C)2 | 1.5E-89 | |||
| 18 | 890 | 66 | 5,404 | Deletion (tandem repeat) | 25.7 | 617 | (G)6 → (G)5 | 3.8E-25 |
| 5,137 | Deletion (tandem repeat | 23.4 | 2555 | (C)3 → (C)2 | 5.0E-10 | |||
| 19 | 274 | 67 | 17,895 | Deletion (tandem repeat) | 25.3 | 617 | (G)6 → (G)5 | 2.8E-68 |
| 16,800 | Deletion (tandem repeat) | 23.8 | 2555 | (C)3 → (C)2 | 4.6E-35 | |||
| 20 | 1516 | 68 | 29,403 | Deletion (tandem repeat) | 24.7 | 617 | (G)6 → (G)5 | 1.2E-87 |
| 28,099 | Deletion (tandem repeat) | 24.1 | 2555 | (C)3 → (C)2 | 2.5E-64 | |||
| 33,567 | SNP (transition) | 28.5 | 1341 | C → G | 0 | |||
| 21 | 1123 | 69 | 11,356 | Deletion (tandem repeat) | 25.6 | 617 | (G)6 → (G)5 | 3.8E-48 |
| 11,187 | Deletion (tandem repeat) | 24.4 | 2555 | (C)3 → (C)2 | 3.2E-31 | |||
| 22 | 2507 | 71 | 51,982 | Deletion (tandem repeat) | 23.3 | 2555 | (C)3 → (C)2 | 2.5E-77 |
| 47,712 | SNP (transition) | 71.9 | 1341 | C → G | 0 | |||
| 23 | 533 | 72 | 43,330 | Deletion (tandem repeat) | 26.0 | 226 | (G)6 → (G)5 | 7.7E-201 |
| 40,426 | Deletion (tandem repeat) | 25.4 | 617 | (G)6 → (G)5 | 1.8E-157 | |||
| 38,886 | Deletion (tandem repeat) | 25.2 | 2555 | (C)3 → (C)2 | 1.9E-141 |
5. Discussion
The emergence of drug resistance in numerous bacteria has significantly heightened the necessity of studying and investigating drug susceptibility in diverse Mycobacterium species. A review of the literature reveals insufficient precise information regarding epidemiology, drug resistance status, and the mechanisms underlying drug resistance in isolates included in studies from Turkey. Our study is based on in vitro data and does not include clinical outcome information or follow-up treatment, as patient records were not accessible. Although demographic factors were not directly analyzed in relation to drug resistance, they were included to provide epidemiological context on the origin and distribution of M. kansasii isolates in Turkey.
The primary purpose of this study was the investigation of the drug susceptibility of forty M. kansasii isolates against nine antibiotics. Furthermore, in this study, we utilized Sanger and Nanopore sequencing to investigate potential mutations in resistant and susceptible strains of M. kansasii and to assess the impact of these gene mutations on their phenotypic characteristics. Given the small size of the regions within the gyrA, gyrB, and rrl genes that play an important role in strain resistance to antibiotics, we employed the first generation of sequencing, Sanger sequencing, to analyze these specific regions.
Since RIF is a crucial first-line drug for treating M. kansasii-related infections, and recent studies on gene mutations associated with RIF resistance in M. kansasii are limited, we employed long-read sequencing technology (Oxford Nanopore Technologies) to sequence the entire rpoB gene, which is approximately 3,600 base pairs in length. Additionally, through a review of previous studies, we found that research has been conducted on the role of the rbpA gene in RIF-resistant M. tuberculosis. However, in the context of RIF-resistant M. kansasii, no prior sequencing of the rbpA gene has been reported. Therefore, in this study, we sequenced the rbpA gene for the first time to investigate whether its mutations contribute to RIF resistance in M. kansasii. The selection of the nine antibiotics evaluated in this study was guided by the CLSI recommendations, which define susceptibility breakpoints for M. kansasii, as well as by their routine use in clinical practice in Turkey (14).
The agents for the treatment of M. kansasii infections, as suggested by the American Thoracic Society, include INH, RIF, and EMB (1, 18). Among these drugs, resistance to rifampicin is critically important as it complicates the treatment process. In this study, there were very high resistance rates for rifampicin (90%). Nonetheless, this rate surpassed the rates observed in other studies. Two studies conducted in Brazil showed 85% and 88% high sensitivity rates to RIF (2, 15). A study from Iran reported 43.5% resistance, while a study from Taiwan showed a 22.6% resistance rate (19). Studies from China revealed two different rates of resistance (20% and 55.4%) (1). A high level of sensitivity was reported from Poland (100%). No resistance to RFB was detected among the forty M. kansasii isolates in our study (3).
In cases of rifampicin resistance, RFB may be utilized as a treatment alternative for infections caused by M. kansasii. Notably, our study found a higher EMB (100.0%) resistance rate compared to Brazil, Taiwan, and European countries such as the United Kingdom and Spain. The rate of EMB resistance in a study conducted in Poland was nearly as high as in Turkey (97%) (3). However, despite the geographical proximity of Iran and Turkey, the rate of drug resistance to EMB in Iran was approximately 48% (19). In our study, all M. kansasii isolates were sensitive (100%) to AMK, CLA, LZD, MXF, and RFB. Other studies have reported similar findings about the effectiveness of AMK, CLA, LZD, MXF, and RFB against M. kansasii (20, 21). Several studies have confirmed the excellent activity of AMK against M. kansasii; only a study from the Netherlands reported resistance to AMK (54%) (22, 23).
AMK susceptibility rates were reported to be 100% in studies conducted in Poland (3), 97% - 100% in Brazil (2, 15), 90% - 92% in China (1, 20), and about 97% in Taiwan (23). Studies on CLA drug resistance against M. kansasii have shown high CLA susceptibility rates in Poland, Taiwan, and Brazil, with only one study in China recording a 20.5% resistance rate against CLR (1). The high resistance rates (90.0%) were observed with CIP in this study. This high rate of resistance to CIP verifies results of studies in Brazil, China, and Taiwan.
Although our study and those from many countries that have researched antibiotic resistance in M. kansasii have shown high sensitivity to LZD, a recent study in Iran reported a drug resistance rate of about 89.1% (19). Moxifloxacin is one of the drugs that can be used as an alternative to first-line drugs in the treatment of infections caused by M. kansasii. Despite the high rate of drug susceptibility reported in Turkey, Poland, and Brazil (100%), recent studies have shown a decrease in susceptibility to this drug in countries such as Iran (65.2%), China (66.1%), and Taiwan (59.5%).
By examining the results of in vitro DST of nine antibiotic agents against M. kansasii and the increasing drug resistance to RIF and EMB in Turkey, the selection of drugs such as AMK, CLA, LZD, MXF, and RFB, which exhibit high sensitivity, can be useful in the treatment of M. kansasii infections. However, it should be noted that the choice of these drugs for treatment may vary across different countries. A limited number of studies exist about the molecular detection of mutations in M. kansasii. Here, we applied partial rpoB, rbpA, gyrA, gyrB, and rrl gene sequencing. The results of the study revealed that 36 M. kansasii isolates resistant to CIP and RIF but sensitive to CLA did not harbor any mutations in the gyrA, gyrB, rbpA, and rrl genes.
Numerous studies have shown that in M. tuberculosis, mutations in the rpoB gene are the main cause of RIF resistance. The rpoB gene in tuberculosis consists of about 3,600 base pairs, and mutations in the RRDR, which consists of 81 base pairs, are known to cause resistance to RIF. Only a few studies have been done on mutations in the RRDR region and the rpoB gene of M. kansasii so far, making it challenging to find a lot of useful information in this area. A study in Poland also could not find any mutations in the gyrA and gyrB genes, but their sequencing of the rrl gene revealed point mutations in A2266C in single M. kansasii isolates (3).
In a study conducted by Klein et al., mutations were identified in RIF-resistant strains at codons 513, 526, and 531 (12). In a more recent study, mutations were recorded in three RIF-resistant strains in the rpoB gene at codons 1249 (417), 1356 (452), 1407 (469), 1479 (493), 1533 (511), and 1536 (512); in the gyrA gene at codons 238, 239 (80), 247, 249 (83), 257, 258 (86), 260, and 261 (87); in the gyrB gene at codons 602, 603 (201), 950 (317), 1332 (444), 1339 (447), 1390, 1392 (464), 1406 (469), and 1594 (532); and in the rrl gene at codons 2058, 2059, and 2266 (19). In this study, we were able to establish SNPs in 6 out of 25 samples in codon 447 and in two samples in codon 457, which are in the RRDR region. These mutations were synonymous and did not cause changes in amino acids.
By sequencing the entire rpoB gene in M. kansasii by ONT, which we did for the first time, we found new mutations outside the RRDR region. The presence of phenotypically resistant isolates in the absence of detectable mutations in the target genes may indicate the involvement of non-genetic factors in antibiotic resistance. Based on previous studies conducted in other mycobacterial species such as M. smegmatis and M. fortuitum, efflux pump systems have been shown to confer resistance by reducing intracellular drug concentrations without the need for specific gene mutations (24, 25). Other contributing factors may include epigenetic regulation or post-transcriptional modifications, the roles of which in M. kansasii drug resistance could be elucidated through transcriptomic and proteomic analyses (26).
Like many scientific studies, our current research has both strengths and limitations. One of the main strengths of this study is the use of long-read Nanopore sequencing technology to investigate mutations in M. kansasii for the first time globally, in combination with Sanger sequencing. Additionally, this is the first comprehensive study from Turkey to evaluate drug resistance in M. kansasii both phenotypically and genotypically.
Despite these strengths, the sample size of 40 isolates may not be sufficient to fully represent the nationwide drug resistance profile of M. kansasii. Another limitation is the lack of investigation into other potential resistance mechanisms, such as efflux pump-related genes or regulatory regions. Additionally, the use of a single reference strain (ATCC 12478) for DST was another limitation. The inclusion of additional resistant reference strains would allow for a more robust interpretation of phenotypic results. In this study, patients’ medical histories and clinical response data were not available. As a result, we were unable to explore potential correlations between specific genetic mutations and clinical outcomes.
Future studies integrating clinical data with molecular findings are essential to better understand the clinical relevance of resistance-associated mutations. Considering the limitations mentioned in this study, future research should include a larger number of isolates collected from various regions of Turkey to gain a more comprehensive understanding of the drug resistance patterns of M. kansasii. Specifically, the high rates of resistance to RIF (90%) and EMB (100%) among the isolates highlight the urgent need for expanded surveillance studies across different regions of Turkey. Additionally, whole genome sequencing (WGS) could be highly effective in identifying alternative resistance mechanisms and novel mutations, particularly those located outside classical resistance-determining regions whose roles in M. kansasii drug resistance have remained unclear to date.
5.1. Conclusions
Our results demonstrate that AMK, CLA, LZD, MXF, and RFB are effective drugs against M. kansasii in vitro. First-line drugs like rifampicin and ethambutol exhibit high resistance rates. Further studies with larger sample sizes are required to monitor drug susceptibility for M. kansasii. The use of new drugs in the treatment of M. kansasii could be a viable option to prevent drug resistance in this species. Additionally, whole genome sequencing of M. kansasii might reveal mechanisms conferring drug resistance.
