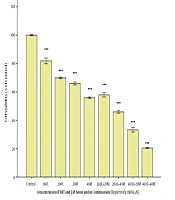
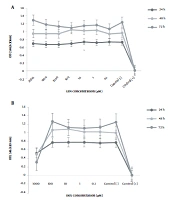
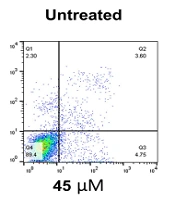
The findings showed that treating the Nalm-6 cell line with mannose alone or in combination with metformin or orlistat decreased viability and induced apoptosis in a concentration-dependent manner. In addition, the expression of GFAT1 and PD-1 genes was reduced.
Recent research has shown the direct inhibitory effect of mannose on the growth of cancer cells both in the cell culture medium and in the oral intake of mannose (
21). Treatment with mannose causes the accumulation of mannose-6-phosphate in the cell, which in turn inhibits glucose metabolism in glycolysis and the Krebs cycle, resulting in a decrease in energy production and inducing endoplasmic reticulum stress and apoptosis (
17,
22).
Mannose alone or in combination with 5-fluorouracil (5-FU) inhibits cell growth in a dose-dependent manner in human colorectal cancer (CRC) cell lines and demonstrated a synergistic effect with 5-FU in all tested cancer cell lines. This study reported decreased dehydrogenase activity of the pentose phosphate pathway, oxidative stress enhancement, and DNA damage induction (
23).
Mannose also reduces the production of NADPH by lowering the isocitrate dehydrogenase 2 (IDH2) level, inhibiting the proliferation of breast cancer cell lines and increasing their sensitivity to oxidant agents. This effect is attributed to the activation of AMPK and the subsequent increase in the expression of E3 ligase, which leads to the destruction of IDH2 (
24). In addition, treatment with mannose leads to defects in glycosylation reactions with the accumulation of mannose 6-phosphate in the cell (
17). Impaired protein glycosylation activated the ATF6 arm of the unfolded protein response (UPR) and subsequent reduction of fatty acid oxidation, accumulation of polyunsaturated fatty acids (PUFA), lipid peroxidation, and cell death through ferroptosis in treatment-resistant AML cells (
25).
In this study, treatment with metformin alone or combined with mannose decreased the viability of the Nalm-6 cell line and decreased GFAT1 gene expression. In addition, metformin, in combination with mannose, decreased PD-1 gene expression.
The reuse of metformin as part of adjuvant therapy for treating several diseases, including various types of cancer, has been widely investigated (
26). Evidence has shown that Slc5A9 carries out the entry of mannose into mammalian cells. The expression of Slc5a9 is induced by metformin (
27,
28). The synergistic effect of metformin with mannose has been shown to correct the synthesis of lipid-linked oligosaccharides and N-glycosylation in Ia cells. This study concluded that AMP-dependent protein kinase may play a role in regulating mannose metabolism (
28). Metformin treatment with cellular metabolic reprogramming inhibited tumor growth by inhibiting mitochondrial oxidative phosphorylation (OXPHOS) in an AMPK-dependent and AMPK-independent manner (
26). The inhibitory effect of metformin on acute lymphoblastic leukemia cells is partially attributed to its inhibitory effect on the mTOR/Akt signaling pathway by AMPK (
29). The study has shown that the activation of AMPK by 2-deoxyglucose, as a mannose analog, (
30) with GFAT1 phosphorylation and its inactivation reduced protein N-glycosylation, induced endoplasmic reticulum stress and subsequently inhibited the growth of pancreatic cancer cells. This effect is synergistically enhanced by metformin through AMPK activation (
22). Since exposure to mannose also activates AMPK (
24), combining metformin with mannose is expected to strengthen each other's effect.
In this study, treatment with orlistat alone or combined with mannose decreased the viability of the Nalm-6 cell line and decreased PD-1 gene expression. In addition, Orlistat, in combination with mannose, reduced the expression of the GFAT1 gene.
An increasing number of studies in recent years have shown that orlistat has an antitumor effect on various cancer cells (
31-
33). Regarding mechanism, orlistat suppresses fatty acid synthase activity and thus reduces free fatty acid production (
34), which also inhibits the activity of fatty acid-binding protein (FABP1) (
35) and interferes with the tumor cell cycle and causes cell death by inducing apoptosis and ferroptosis (
36).
Orlistat can reduce palmitoylation and activation of AKT and prevent liver tumorigenesis by limiting palmitic acid (PA) synthesis or suppressing AKT modification (
37). Fatty acid synthase (FASN) is highly expressed in leukemic cancer, and its inhibition impairs angiogenesis (
38,
39). Orlistat has been shown to decrease PDL-1 and PD-1 levels significantly, and the decrease in PDL-1 levels is associated with FASN inhibition (
40).
In the present study, a decrease was observed in the expression of GFAT1. GFAT1 expression is often increased in cancer cells (
41). Inhibiting the hexosamine pathway reduces the production of UDP-GlcNAc and glycosylation reactions. In this regard, inhibition of GFAT1 leads to endoplasmic reticulum-related apoptosis by disrupting protein N-glycosylation in pancreatic cancer cells (
41). According to some studies, including a survey conducted on lung cancer cells, inhibition of GFAT1 decreases the stability of the PD-L1 protein (
42).
In this study, a decrease was also observed in
PD-1 gene expression. Programmed cell death protein 1 is an N-glycosylated protein, and the glycosylation process plays a role in determining its stability and expression on the surface of T lymphocytes and its interaction with the corresponding ligand, PD-L1. As lymphocytes and tumor cells activate, glycosylation pathways become more active (
25). In this regard, 2-deoxy-D-glucose, as a mannose analogue, can inhibit the biosynthesis of N-glycosylated compounds in the pancreatic adenocarcinoma tumor model and strengthen immunotherapy (
30). Overall, this study showed that treatment with mannose, metformin, and orlistat decreases cell survival, induces apoptosis in Nalam-6 cells, and decreases the expression of GFAT1 and
PD-1 genes. In addition, combining mannose with metformin and mannose with orlistat synergistically inhibited cell growth.
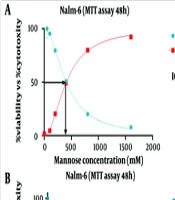



. The data are from three independent trials.")
-preview.webp "The synergistic effect of mannose compounds (Man + Met and Man + Orl) on inhibiting the growth of the Nalm-6 cell line. The Combination Index (CI) was determined via an isobologram test based on the Chou-Talalay approach. Combination Index < 1, synergistic effect; CI = 1, additive effect; CI > 1, antagonistic effect. Data depicted are after three independent trials. The green line in the polygonogram shows the effect of synergy at the combined concentration of Fa = 0.5. Man, Mannose; Orl, orlistat; Met, metformin.")
.")