According to the results of this study, the VPA-offspring rats displayed autistic-like behaviors, such as increased pain tolerance, anxiety-like behaviors, and imbalance. A previous study found that the brains of rats that received VPA during fetal life had developed abnormalities, such as anatomical changes in the cerebellum, amygdala, hippocampus, frontal lobes, and parietal lobes, similar to those seen in children with autism. Therefore, the VPA model has both structural and behavioral characteristics similar to those in autism, and therefore, it is a suitable model for the assessment of autistic-like behaviors in rodents (
24).
On the other hand, administering vitamins D and E for four weeks significantly improved behavioral disorders and oxidative stress markers. One common finding between different behavioral and oxidative tests in this study was that the administration of vitamins D and E, or combined, produced their effects only in rats that received VPA but not in healthy control groups. The positive effects of these vitamins have been studied so far for other disorders related to the nervous system, such as chronic stress-induced depressive-like behaviors and vitamin D's significant effect on alleviating anhedonia. This effect is strongly suggested to be related to the regulation of vitamin D on neurotransmitter systems, especially dopamine and reward systems (
26,
27). Likewise, another study has explored vitamin E deficiency during development and its association with glutamate levels and anxiety related to familial ataxia in adult mice. Vitamin E supplements alleviated anxiety symptoms (
28). In relation to the autism spectrum, other studies have provided evidence of vitamin E's relieving effect on VPA-induced autistic-like behaviors, such as anxiety and repetitive/stereotypic-like behaviors, and sociability impairments in rats exposed to VPA before birth (
29). Also, the administration of vitamin D supplements markedly relieved repetitive behaviors and increased grooming in rat offspring treated with VPA during pregnancy (
30).
In the present study, rat offspring with maternal VPA treatment displayed significantly higher pain thresholds than normal rats. Along with our result, another study also reported an increase in the pain threshold in offspring rats caused by maternal VPA treatment, which was decreased using pregabalin (
31). It seems that under the influence of VPA, the nitric oxide (NO) signaling pathway is probably affected by imbalances in neurotransmitter release, immune disorders, and oxidative stress (
32). Since pain sensation in the periphery depends on the presence of different local inflammatory mediators, including NO (
33), therefore, an imbalance in NO production alters the threshold of pain sensation as it happens in autistic patients (
34).
In this, as well as some other studies, it has been indicated that there is a relation between vitamin D and NO function, specifically in relation to neural diseases such as autism spectrum. NO directly regulate vitamin D expression through
CYP27B1. On the other hand, since inflammatory factors increase the expression of inducible Nitric Oxide synthase (iNOS) and, consequently, produce NO, vitamin D modulates the inflammatory processes and reduces NO production (
32).
In the present study, using rotarod, EPM, OF, and TM that measured the rat's motor balance, anxiety levels, and spatial learning/memory indicated that VPA had induced imbalance and mal-coordination, increased anxiety, and decreased level of spatial learning in the rat. Previous studies indicate that autism spectrum and animal models such as VPA-induced autistic-like behaviors produce motor deficits through damage to the cerebellum and its connections to the motor and prefrontal cortex (
35). These damages are a possible reason for mal-coordination in humans and animals under a model of autistic disorders. In line with our findings, Edalatmanesh and Samimi study showed that a decrease in balance and learning in rats with the VPA model of autism indicated a probable decrease in BDNF levels or damage to Purkinje cells in the cerebellum. They used gallic acid as an antioxidant to alleviate motor disorders (
36).
It is mostly indicated that the underlying reasons for motor deficiencies related to autistic syndrome are immune system malfunction, oxidative stress, and neurotransmitter imbalance (
35). Studies have shown that patients with autism have higher plasma levels of autoimmune indicators such as anti-nuclear antibodies as well as pro-inflammatory cytokines such as IL-6 and tumor necrosis factor-alpha (TNF-α) in different tissues, which may be associated with cognitive impairment in these patients. An increase in oxidative stress and mitochondrial dysfunction can produce neural damage. Also, autistic disorder is associated with a deficiency in GABA neurotransmitters as well as serotonin and dopamine (
37).
Several reports also show that autism and the VPA model of autistic disorder increase excitability and anxiety in humans and animals (
31,
38). Some relate the anxiety to hyperactivation of the amygdala due to alteration in GABAergic transmission or increased expression of GluN2A- and GluN2B-NMDA receptors and amplified long-term potentiation (LTP) and increased excitatory synapses in the cortex (
38). Some studies suggest that the reason behind the delayed development of the sensory system, as well as anxiety and spatial learning in autistic-like syndromes, is related to an imbalance in GABA production and release in different parts of the cortex as well as the hippocampus and cerebellum. Using a VPA model of disease, they revealed decreased expression of glutamic acid decarboxylase 67, GAD67, expression in the hippocampus and cerebellum from adolescence to adulthood and temporal cortex in adulthood, while increased expression was found in the prefrontal cortex from adolescence to adulthood (
39). Still, other studies are focusing on oxidation stress and immune system dysfunction as potential reasons for autistic behaviors, such as anxiety and learning deficits (
40).
In our study, administering vitamins D and E effectively improved rat function in rotarod, anxiety, and spatial learning tests. Vitamins D and E have powerful antioxidants (
28,
37), anti-inflammatory (
41), and neurotransmitter regulatory effects (
26,
37). For the GABA inhibitory system, vitamin E creates a balance in activates of the enzymes converting glutamate to GABA in different brain regions (
42); also, mice with vitamin D deficiency have shown a large decrease in GAD 65/67 expression in the whole brain. The balance in the activity of the GABA inhibitory system has an important effect on the neurogenesis and development of the brain (
43). Therefore, it can be strongly suggested that the improvement in movement and balance, as well as anxiety and spatial learning deficits observed in the VPA-induced autistic behaviors, are related to both vitamins, and since the action of each vitamin has been some overlapped with another (like both acting as antioxidants), maybe with a different mechanism of action, that might be the reason for the lack of augmented response seen, when both were administered together.
Our findings in the present study indicated that the levels of GLU and CAT were decreased, and the level of MDA was increased in the brains of VPA-offspring, while the administration of vitamins D and E decreased the brain levels of MDA. Previous studies indicate that increased pro-oxidants such as NO and decreased antioxidants such as GLU can cause oxidative stress. Increased oxidative stress leads to membrane lipid defects, mitochondrial dysfunction, inflammation, and immunodeficiency in children with autism, resulting in behavioral, sleep, and gastrointestinal disorders. On the other hand, clinical trials documented the improvement in the behaviors of children with autism undergoing treatment with antioxidants (
6). Similarly, in another study, administrating piperin as an antioxidant agent adjusted the levels of the oxidative factor, MDA, and antioxidative agents CAT and GLU in a model of VPA-induced autism (
44). Ghanizadeh et al. investigated various factors affecting the oxidative stress level that increased and decreased GLU and SOD levels in children with autism. They also suggested antioxidants as important factors in the treatment of autism (
45).
Since oxidative stress is one of the major factors in the pathophysiology of autism, there have been various studies investigating the effects of different types of antioxidants as treatment options. Among all, vitamins, specifically vitamin D and E, have received special interest recently; there have been a good number of studies investigating their antioxidative effects under conditions that markedly raise oxidative stress incidents, such as chronic psychological stress. Under a model of chronic mild stress, a marked increase in the pro-inflammatory factors happened, along with a significant reduction in the SOD and glutathione peroxidase (GPx) as indigenous antioxidants. Administration of vitamin D reduced the pro-inflammatory cytokines and raised the suppressed levels of SOD and GPx (
41). Likewise, Kocovska et al. investigated the association between vitamin D and autism by reviewing 15 years of animal, molecular, cellular and physiological, post-mortem brain, and neuronal imaging studies related to the role of vitamin D in autism and suggested that patients with autism are likely to be exposed to vitamin D deficiency, which might have raised the odds ratio of autistic behaviors along with other factors (
18). On the other hand, Gumpricht and Rockway investigated the role of omega-3 fatty acids and Tocotrienol-rich vitamin E in reducing symptoms of Alzheimer's disease and neurodevelopmental disorders, such as autism. Their results showed an increase in the level of oxidative stress and a change in antioxidant defense factors and neuroinflammation in autistic children. Their study suggested that omega-3 fatty acids and vitamin E could be used as supplements to alleviate behavioral and cognitive symptoms in neurodevelopmental and neurodegenerative disorders such as autism and Alzheimer's disease (
16). Also, another study in 2010 described the role of Thai brown rice in reducing oxidative stress, and the results showed that it reduced MDA levels as a marker of oxidative stress. Thai brown rice contains high levels of vitamins D and E (
46).
This study, for the first time, investigated the effects of both vitamins D and E as strong antioxidants and anti-inflammatory agents in alleviating some disturbing syndromes related to maternal VPA administration in male offspring, such as an increase in pain threshold (indicating a high risk of tissue damage because of decreased levels of sense of pain), motor balance incoordination, anxiety, and learning deficiencies. By referring to previous studies, indicated effects may also be implied for human beings. Further investigation strongly suggests understanding the mechanism underlying their actions.
5.1. Conclusion
In this study, the model of maternal VPA-induced autistic-like behaviors in the rat male offspring; elevated pain threshold, produced motor imbalance, increased anxiety, and diminished spatial learning. The goal of this study was to evaluate the effect of vitamins D and E on the alleviation of maternal VPA-induced syndromes, as well as the levels of oxidative stress in offspring. Both vitamins, individually or in combination, regulated the rat response to thermal pain, motor balance, levels of anxiety, and spatial learning in rat offspring. This is the first study showing the effects of both vitamins D and E, as strong antioxidants, on sensory, motor, mood, and cognitive deficiencies and oxidative stress induced by maternal VPA consumption in offspring. This study also confirmed this suggestion that using both vitamins (perhaps as supplements) doesn't have an augmented effect in relieving the deficiencies.
5.2. Limitations
The main limitation of this study was the lack of accessibility to most of the tools and devices used for autistic-like behavior studies, like those that are used for investigating the social interaction deficit.








.")
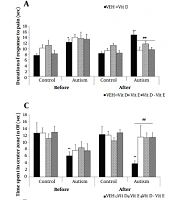
![Effects of vitamin D E and combined on: A, Pain responses by a hot plate; B, Motor balance by rotarod; C, Anxiety-like behavior by an open field; D, Anxiety-like behavior, by the elevated maze; and E, Spatial learning, by T-maze, in male offspring rats that prenatally exposed to VPA [data are expressed as mean ± SEM; n = 6; **P < 0.01, *P < 0.05 versus control-VEH (vehicle) group; ##P < 0.01 versus VPA-VEH group (autism); &&P < 0.01 post-treatment versus pre-treatment with vitamins; VEH, vehicle].](https://brieflands.com/journals/mejrh/articles/129627/figures/mejrh-129627-i002-F2-preview.webp "Effects of vitamin D E and combined on: A, Pain responses by a hot plate; B, Motor balance by rotarod; C, Anxiety-like behavior by an open field; D, Anxiety-like behavior, by the elevated maze; and E, Spatial learning, by T-maze, in male offspring rats that prenatally exposed to VPA [data are expressed as mean ± SEM; n = 6; **P < 0.01, *P < 0.05 versus control-VEH (vehicle) group; ##P < 0.01 versus VPA-VEH group (autism); &&P < 0.01 post-treatment versus pre-treatment with vitamins; VEH, vehicle].")
![Effects of vitamin D, E, and combined on brain stress oxidative markers. A, malondialdehyde; B, glutathione; C, catalase in male offspring rats that were prenatally exposed to VPA [data are expressed as mean ± SEM; n = 6; **P < 0.01; *P < 0.05 versus control-VEH; ##P < 0.01; #P < 0.05 versus VPA (autism); VEH, vehicle].](https://brieflands.com/journals/mejrh/articles/129627/figures/mejrh-129627-i003-F3-preview.webp "Effects of vitamin D, E, and combined on brain stress oxidative markers. A, malondialdehyde; B, glutathione; C, catalase in male offspring rats that were prenatally exposed to VPA [data are expressed as mean ± SEM; n = 6; **P < 0.01; *P < 0.05 versus control-VEH; ##P < 0.01; #P < 0.05 versus VPA (autism); VEH, vehicle].")