




1. Background
2. Objectives
3. Methods
3.1. Laboratory Animals and Rearing House
3.2. Animal Grouping
3.3. Production Method of Nano-selenium
3.4. Method of Preparing and Studying Slides of Testicular Tissue

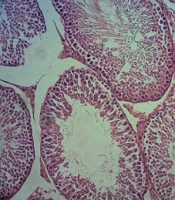
Photomicrograph of testicular tissue in a control group of young rat; magnification 100×, hematoxylin-eosin staining

Photomicrograph of testicular tissue in a control group of old rat; magnification 100×, hematoxylin-eosin staining

Photomicrograph of testicular tissue in the group receiving 0.2 mg/kg BW nano-selenium of old rat; magnification 100×, hematoxylin-eosin staining

Photomicrograph of testicular tissue in the group receiving 0.4 mg/kg BW nano-selenium of old rat; magnification 100×, hematoxylin-eosin staining
3.5. Ethical Consideration
3.6. Statistical Analysis Method
4. Results
4.1. Number of Spermatogonia and Spermatocyte
 in different weeks")
Comparison of the nano-selenium effect on changes in the number of spermatogonia, spermatocytes, spermatids, and spermatozoid (×106) in different weeks
| Treatment | Spermatogonia | Spermatocyte | Spermatid | Spermatozoid | ||||
|---|---|---|---|---|---|---|---|---|
| Young | Old | Young | Old | Young | Old | Young | Old | |
| Control | ||||||||
| Week 1 | 63.3 | 69.3 | 105.0 | 36.7 | 125.0 | 26.7 | 165.0 | 21.7 |
| Week 2 | 75.0 | 62.7 | 136.7 | 113.3 | 164.0 | 43.3 | 190.0 | 40.0 |
| Week 3 | 67.0 | 65.0 | 121.7 | 131.7 | 132.7 | 53.3 | 193.3 | 70.0 |
| Week 4 | 64.0 | 83.3 | 132.3 | 113.3 | 123.7 | 40.0 | 173.3 | 43.3 |
| Placebo | ||||||||
| Week 1 | 76.7 | 55.0 | 118.3 | 50.0 | 143.3 | 40.0 | 175.0 | 38.3 |
| Week 2 | 70.0 | 63.3 | 130.0 | 86.0 | 153.3 | 36.7 | 185.0 | 60.0 |
| Week 3 | 65.7 | 63.3 | 186.0 | 106.7 | 184.7 | 36.7 | 259.3 | 45.0 |
| Week 4 | 70.3 | 80.0 | 141.7 | 103.3 | 149.0 | 23.3 | 165.0 | 40.0 |
| 0.2 | ||||||||
| Week 1 | 70.0 | 64.7 | 133.3 | 157.0 | 145.0 | 96.3 | 171.7 | 91.3 |
| Week 2 | 86.7 | 58.3 | 103.3 | 146.7 | 133.3 | 143.3 | 190.0 | 146.7 |
| Week 3 | 74.7 | 76.7 | 193.0 | 126.7 | 213.0 | 56.7 | 285.3 | 80.0 |
| Week 4 | 67.0 | 80.0 | 180.0 | 116.7 | 165.3 | 60.0 | 193.3 | 66.7 |
| 0.4 | ||||||||
| Week 1 | 73.3 | 73.3 | 153.3 | 90.0 | 163.3 | 123.3 | 181.7 | 143.3 |
| Week 2 | 65.0 | 78.3 | 113.3 | 136.7 | 126.7 | 146.7 | 133.3 | 176.7 |
| Week 3 | 70.0 | 71.7 | 176.0 | 115.0 | 209.7 | 93.3 | 277.0 | 86.7 |
| Week 4 | 71.0 | 90.0 | 158.3 | 123.3 | 180.0 | 90.0 | 250.0 | 93.3 |
| P-value | ||||||||
| Treatment | 0.90 | 0.05 | 0.002 | 0.024 | ||||
| Age | 0.36 | 0.001 | 0.00 | 0.00 | ||||
| Week | 0.23 | 0.03 | 0.49 | 0.21 | ||||
| Treatment × age | 0.26 | 0.40 | 0.11 | 0.37 | ||||
| Number/Groups | Young | Old | RCTA | Probability Level | |
|---|---|---|---|---|---|
| Young | Old | ||||
| Spermatogonia | 0.437 | 0.410 | |||
| Control | 123.92 | 86.50 | 69.8 | ||
| Placebo | 144.00 | 98.80 | 68.61 | ||
| 0.2 | 152.40 | 136.75 | 89.73 | ||
| 0.4 | 150.30 | 116.25 | 77.35 | ||
| Spermatocytes | 0.313 | 0.090 | |||
| Control | 113.90 | 98.65 | 86.61 | ||
| Placebo | 142.40 | 86.50 | 60.74 | ||
| 0.2 | 144.00 | 106.75 | 74.13 | ||
| 0.4 | 150.30 | 116.25 | 77.35 | ||
| Spermatids | 0.263 | 0.001 | |||
| Control | 136.33 | 40.83 B | 29.95 | ||
| Placebo | 157.58 | 34.17 B | 21.68 | ||
| 0.2 | 164.20 | 89.10 A | 54.26 | ||
| 0.4 | 169.90 | 113.30 A | 66.69 | ||
| Spermatozoid | 0.544 | 0.0004 | |||
| Control | 180.42 | 43.75 B | 24.25 | ||
| Placebo | 196.10 | 45.83 B | 23.37 | ||
| 0.2 | 210.10 | 96.20 A | 45.79 | ||
| 0.4 | 210.50 | 125.00 A | 59.38 | ||
Abbreviation: RCTA, the ratio of changes between the two ages.
a Columns with at least one common capital letter (A and B) in superscript do not differ significantly.
4.2. Number of Spermatid

Changes in the use of nano-selenium on the number spermatogonia, spermatocyte, spermatid, and Spermatozoid in old rats
4.3. Number of Spermatozoid
| Treatment | S (×106)/mL | LS (%) | DS (%) | LSD (%) | DDS (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Young | Old | Young | Old | Young | Old | Young | Old | Young | Old | |
| Control | 180.42 B | 43.75 | 81.50 | 57.00 | 10.50 | 31.00 | 6.00 | 6.50 | 2.00 | 5.50 |
| Placebo | 196.10 B | 45.83 | 74.25 | 62.50 | 20.75 | 23.00 | 2.50 | 10.00 | 2.50 | 4.50 |
| 0.2 | 210.10 A | 107.94 | 76.00 | 82.75 | 18.00 | 14.25 | 4.00 | 2.50 | 2.00 | 0.50 |
| 0.4 | 210.50 A | 125.00 | 79.00 | 88.25 | 10.00 | 7.00 | 9.25 | 3.50 | 1.75 | 1.25 |
Abbreviations: S, spermatozoa; LS, live spermatozoa; DS, dead spermatozoa; LSD, live spermatozoa damaged; DDS, dead damaged spermatozoa.
a Columns with at least one common capital letter (A and B) in superscript do not differ significantly.
| Cells | Spermatogonia | Spermatocytes | Spermatid |
|---|---|---|---|
| Spermatocytes | -0.153 (0.418) | ||
| Spermatid | -0.070 (0.713) | 0.708 (0.000) | |
| Spermatozoid | -0.052 (0.783) | 0.685 (0.000) | 0.968 (0.000) |
a The numbers in parentheses indicated different statistical levels.
4.4. Number of Sertoli and Leydig
| Groups | Young | Old | Probability level | |
|---|---|---|---|---|
| Young | Old | |||
| Sertoli | 0.44 | 0.26 | ||
| Control | 19.00 | 13.00 | ||
| Placebo | 19.00 | 14.00 | ||
| 0.2 | 20.00 | 14.00 | ||
| 0.4 | 20.00 | 14.00 | ||
| Leydig | 0.31 | 0.25 | ||
| Control | 17.00 | 17.00 | ||
| Placebo | 18.00 | 17.00 | ||
| 0.2 | 17.00 | 18.00 | ||
| 0.4 | 18.00 | 17.00 | ||
4.5. Correlation and Regression Equations
")
Graphs and regression equations between the number of spermatogonia, spermatocytes, spermatids, and spermatozoid (×106)