This study was carried out for the first time on a rare and endemic species of Algerian flora: Origanum floribundum (Lamiaceae family). Moreover, pharmacological studies on this species are very limited. We are trying to evaluate the hepatoprotective effect of lyophilized hydro-ethanolic extract of O. floribundum and to understand its molecular mechanisms on an acute model of hepatoxicity induced by a single toxic dose of paracetamol (2 g/kg).
Currently, scientists have a strong interest in naturally occurring secondary metabolites. They have examined their antioxidant properties that may be the treatment of various ailments due to their health benefits (
30). In terms of their functions, the antioxidants can be classified into scavenger antioxidants, preventive antioxidants and
de novo and repair antioxidants (
31). Consequently, the antioxidant activity of
O. floribundum extract was carried out using four different assays under different conditions: the capacity of hydroethanolic extract for scavenging free radicals was assessed by two methods: DPPH free radical scavenging and ABTS
°+ cation radical discoloration. Assessment by reduction of cupric (Cu II) to their lower valiancy state known as the CUPRAC method was used. Finally, the reducing power assay was carried out.
The effect of antioxidants on DPPH or ABTS radicals scavenging is achieved by hydrogen or electron transfer followed by proton transfer to become stable molecules (
31). The reaction of antioxidants with DPPH or ABTS
°+ is followed by a decrease in their absorption at 517 and 734nm respectively. The relative antioxidant capacity for scavenging radicals is expressed by inhibitory concentration (IC
50), which corresponds to the concentration of the extract to obtain a 50% reduction in the initial free radical (DPPH or ABTS) concentration. A higher radical capture activity is associated with lower IC
50 levels. Following our results, the hydro-ethanolic extract of
O. floribundum has a potent antioxidant. These results are related to the high content of phenolic compounds. Also, it is known that phenolic compounds act mainly as (primary) scavenger antioxidants of free radicals.
In the literature, a powerful radical scavenging antioxidant usually functions as a powerful reducer (
31). The CUPRAC method is based on the reaction between complexes of probes (Nc) with the reduced form of the metals giving visible absorption bands with maximum intensities at 450nm (
32). The reductive potential is associated with antioxidant activity. The reducing power property of an extract indicates the capacity to act as an electron donor by which the lipid peroxidation process can be stopped by reducing oxidative products to progressively stable substances (
33). In this study, the
O. floribundum extract gave an absorbance value 02 folds higher than that expressed by BHA standard (A
0, 5 values: 16, 64 ± 0, 9 and 8, 41 ± 0, 67 µg/ml respectively) that means, the reducing power of the standard antioxidant is more than twice as high as that of the extract tested. Therefore, the extract has a powerful reducing power.
AC induced liver injury and has served as the most important model, useful for testing the hepatoprotective activity of medicinal plants (
34). After 24 h of AC intoxicated rats, abnormal liver indicators become apparent in serum include by elevated serum aminotransferases activities (AST, ALT) and ALP. Since these enzymes are exclusively intracellular, their release is explained by the loss of integrity of the hepatocytes’ membranes and lipids’ peroxidations (
35,
36). These findings are confirmed after the observation of histological sections that demonstrate massive necrosis caused by AC. Furthermore, significant reductions in serum protein and albumin levels were marked. These results are related to the loss of functional integrity of the liver and a reduction in the number of hepatocytes through necrosis (
30,
37). The animals pre-treated with
O. floribundum (400 m/kg) revealed significant protection of liver tissue against AC that appeared in the normalization of serum levels of liver function markers (
Table 2) and was confirmed after observation of histological sections (
Figure 1). These results concluded that
O. floribundum has a promising hepatoprotective effect.
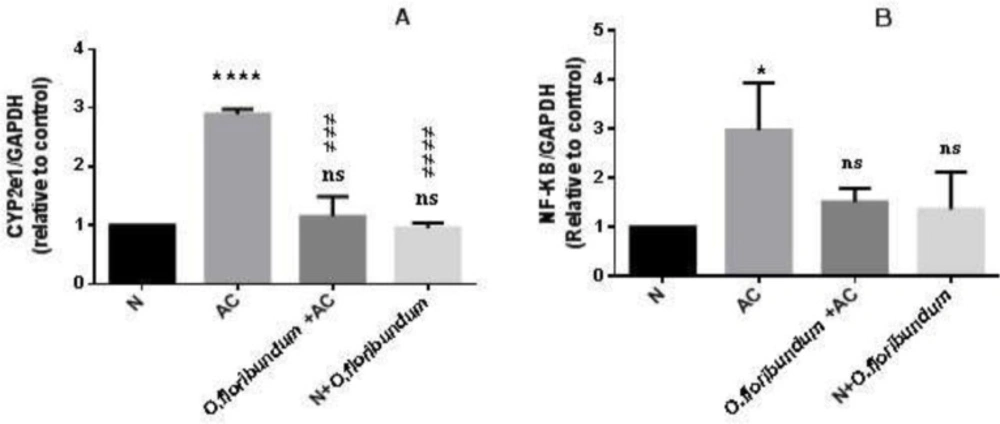
AC-induced hepatotoxicity is conducted by the action of cytochrome P 450 mainly CYP2E1 to form reactive metabolites: N-acetyl p-benzoquinone (NAPQI) and releases a significant amount of ROS. High levels of NAPQI induce a decline in the GSH stores in cytosolic and mitochondrial levels (
5). The decrease in mitochondrial GSH levels is explained by the presence of CYP2E1 also into mitochondria (
38) and in this way, NAPQI is generated within to mitochondria. GSH depletion is the main event that initiates lipid peroxidation and protein adducts formation especially in the mitochondrial respiratory chain. Consequently, it promotes overproduction of ROS and selective oxidant stress. This aggravates paracetamol-induced hepatotoxicity.
Since cytochrome P450 particularly CYP2E1, is the main oxidation system of paracetamol into NAPQI and ROS that triggers a cascade of the process resulting in hepatotoxicity. Also, the overexpression of the cytochorome P450
CYP2E1gene in AC overdose is confirmed by several studies (
39,
40). We investigated the effect of
O.floribundum extract on the expression of the
CYP2E1 gene. As expected, in the present study, AC overdose induces
CYP2E1 gene expression very significantly. However, pre-treatments with
O. floribundum extract normalize
CYP2E1 gene expression to a level similar to the normal group. Interestingly, this finding supported for the first time that
O.floribundum has a molecular protection mechanism against AC through downregulation of
CYP2E1 gene expression. This result means that
O.floribundum extract suppresses the conversion of AC into NAPQI and free radicals in the liver and thereby could explain the improvement of GSH level and redox status. Therefore, one of the molecular mechanisms of the protection of
O. floribundum against paracetamol-induced hepatotoxicity is interaction with the expression of
CYP2E1 gene.
Several previous reports exhibit that polyphenols may be effective in regulating the cytochrome CYP450 system via the inhibiting of their activity, the decline in their liver content and their expression (
41). For example, rosmarinic acid caused marked inhibition of the CYP2E1 enzymatic activity in AC overdose (
42). This phenolic acid is an excellent chemotaxonomic marker present in the Nepetoideae subfamily where our study plant belongs:
Origanum floribundum. However, according to the literature review, this phenolic acid is not yet confirmed in the phenolic composition of
O. floribundum. NF_kB is a transcriptional factor related to stress-induced inflammation (
43). In this study, we found several markers of inflammation at the tissue level in the AC intoxicated group, such as severe congestion, sinusoid dilation, and inflammatory cell infiltration. These results are in agreement with several previous studies, which often linked inflammatory response with AC-induced liver damage (
5,
44). However, treatment with
O. floribundum extract declines all these events. The above data indicate that activation of the NF_κB signaling pathway is linked to inflammatory response and cellular dysfunction and damage (
43). In the present study, AC intoxication resulted in over-expression of the
NF_κB gene, which is in concurrence with (
45), who detected strong phosphorylated protein expressions of NF_κB in AC overdose. Interestingly, in the present work, we demonstrated for the first time that
O. floribundum reduced
NF_κB gene expression-induced hepatic inflammation in rats. In a previous study,
Origanum majorana extract was revealed to suppress
NF_κB activation against breast cancer via anti-metastatic activity (
46). Similarly, this study demonstrated that
O.floribundum significantly attenuated liver injury in rat partly via the inhibition of
NF_κB gene expression.NF_κB signaling pathway activation in AC overdose caused the transcription of several inflammation-associated genes (
TNF-α, IL-1B,IL- 6) (
5,
43). It, therefore, seems that the inhibitory effect on the expression of the
NF_κB gene may be the result of the anti-inflammatory effects of
O. floribundum.
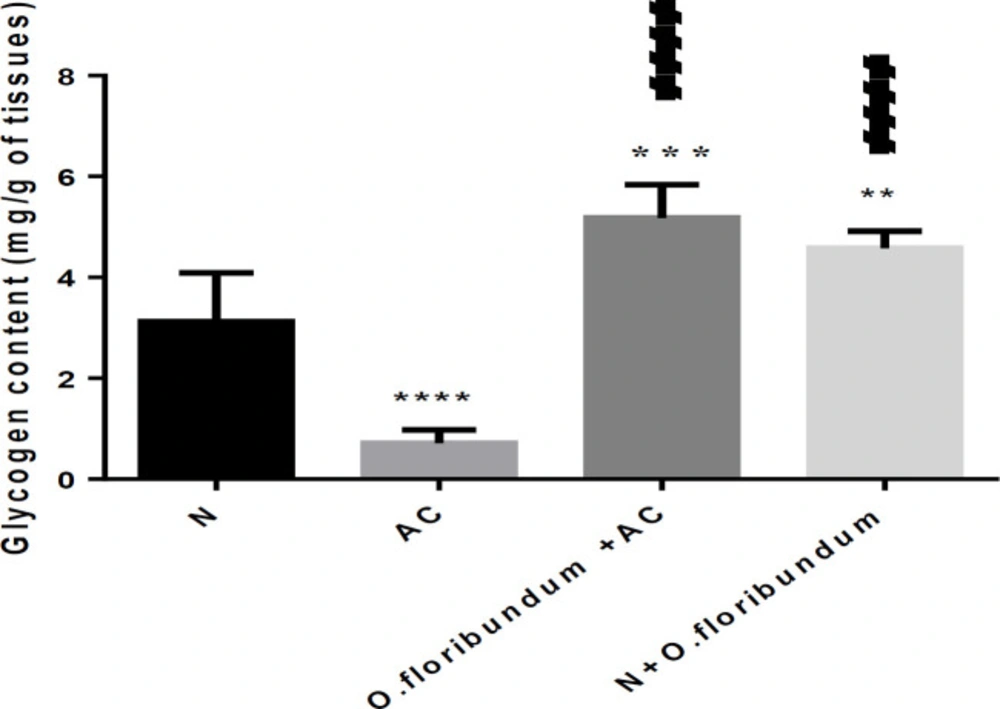
Among the specific functions of the liver is the storage of glycogen. In this study, AC intoxication resulted in depletion of liver glycogen which is in concurrence with many research (
47,
48). Indeed, in the case of paracetamol intoxication, several authors confirm the activation of glycogen degradation via the glycogenolysis pathway (
49) by activating the key enzyme of glycogenolysis: glycogen phosphorylase A (
50). In fact, the main pathways of AC metabolism are conjugation through glucuronosyl-transferase and sulfo-transferase to produce no toxic metabolites. These routes depend on the accessibility of their co-substrates: UDP-glucuronic acid (UDPGA) and 3’-phosphoadenosine5’-phosphate (PAPS), respectively. The latter is provided by glycogenolysis (
51). On the other hand, the glycogen level in the animals intoxicated and pre-treated by
O. floribundum is significantly high compared to the normal group. We suggest, in this case by the effect of the bio-molecules present in the extract of the plant to increase a certain growth under the effect of the toxic and resist the damage caused by AC through prevention of glycogenolysis.
From the results found above, it can be explained that paracetamol overdose increased oxidative stress and mitochondrial dysfunction. Indeed, depletion of GSH and elevation of MDA end products both in cytosolic and mitochondrial levels were recorded. Moreover, we registered a decrease in catalase activities both in cytosol and mitochondria and glutathione S-transferase activities in the cytosol.
The protective effect of
O.floribundum could be associated with its ability to prevent the increase of oxidative damage and decrease of antioxidant enzyme activity. Interestingly, we have found that
O. floribundum increased the normal level of cytosolic GSH significantly. Well, these results are partly due to the high contents of
O.floribundum in phenolic and flavonoïds compounds, secondary metabolites known for their antioxidant power. This later has been proven in this study by several different tests “
in vitro”. In addition, there are some studies on the effects of particular flavonoids on glutathione levels. Moskaug has been found that quercetin increases the expression of γ-glutamylcysteine synthetase with a concomitant increase in the intracellular glutathione (
52). A preliminary analysis (Result not yet published) by chromatographic separation coupled with mass spectrometry (GC-MS) shows that the extract of
O. floribundum is very rich in bioactive molecules (52 compounds), including fatty acids (n-Hexadecanoic acid), catechol and, indole. A derivative of indole has been proven by Park (
53) to be able to combine directly with NAPQI. According to these results and for the first time,
O.floribundum treatment can increase GSH content and induce both enzymatic and no enzymatic antioxidant capacity.
Mitochondrion plays a key function in cells survival. Indeed, it is responsible for 95% of cellular energy needs. In this study, we recorded mitochondrial dysfunction through depletion of glycogen, stress oxidative and, mitochondrial swelling. On the other hand, O.floribundum pretreatment demonstrates a protective effect on AC-induced mitochondrial dysfunction. It is well established that NAPQI overproduction inset the mitochondria block the respiratory chain, increases cell calcium, the formation of ROS including peroxynitrite and, activation of the mitochondrial permeability transition leading to hepatocyte necrosis. The protective effect of O. floribundum on AC-induced mitochondrial dysfunction could be associated with its antioxidant effects.
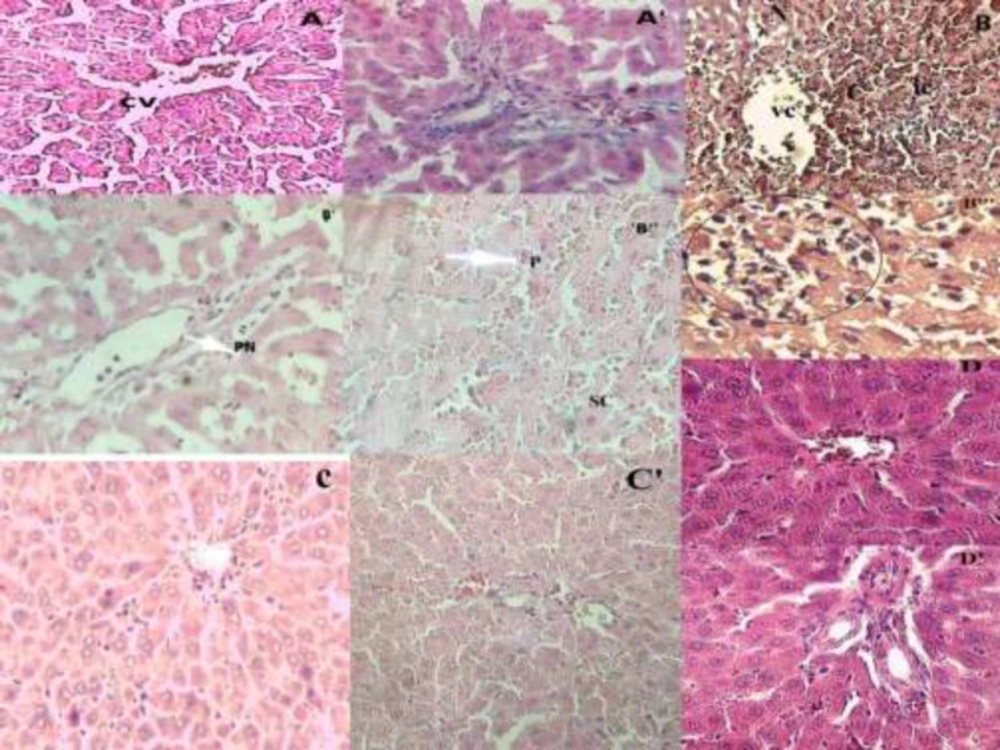
Effect of O. floribundum extract on liver histological changes following AC toxicity in rats: (A) control group showing the normal structure of the liver, (A') normal portal space of control group with veins and arteries of regular structure; (B, B", B'"): AC treated groups with massive Porto-centrilobular necrosis. Swollen hepatocytes (B") hepatocyte with nuclear pyknosis (B") inflammatory cells infiltration for (B'’’), B’: portal space of AC treated group with piecemeal necrosis, Figure C: O. floribundum +AC pretreated group showing sub-normal architecture, Figure C’: normal portal space of O floribundum+AC pretreated group Figure D: normal liver for O. floribundum pretreated group, Figure D’: normal portal space of O. floribundum group (Hematoxylin Eosin X200); X400 for figure B”’. CV: centrilobulare vein; C: congestion; P: pyknos; PN: pieacemeal necrosis; N: necrosis; SC: swelling cell, IC: inflammatory cells
(A and B) Study of the CYP2E1 and NF-KB genes expression by qRT-PCR in the liver rat
effect of O. floribundum on liver glycogen content in different treated groups
| Extract | Antioxidant activity |
|---|
| DPPH assay | ABTS assay | CUPRAC assay | Reducing power assay |
|---|
| CI50 (µg/mL) | CI50 (µg/mL) | A0,5 (µg/mL) | A0.5 (µg/mL) |
|---|
| Extract of O.floribundum | 15,69 ± 1,00 | 7,15 ± 0, 37 | 18, 41 ± 0, 81 | 16, 64 ± 0, 9 |
| BHTa | 22, 32 ± 1, 19 | 1,59 ± 0,03 | 9, 62 ± 0, 87 | >50 |
| BHAa | 5, 73 ± 0, 41 | 1,03 ± 0,0 | 3, 64 ± 0,19 | 8, 41 ± 0, 67 |
| Ascorbic Acida | Nt | Nt | Nt | 9, 01 ± 1, 46 |
| Groups | ALT(IU/L) | AST(IU/L) | ALP(IU/L) | Albumin(g/L) | Protein(g/L) |
|---|
| N | 68,21 ± 17,53 | 180,24 ± 51,75 | 119,33 ± 28, 44 | 40,38 ± 1,25 | 78,93 ± 2,74 |
| AC | 1384,13 ± 197,58**** | 2636,71 ± 243,69**** | 200,66 ± 62,36* | 37,19 ± 0, 46* | 73,08 ± 1,46** |
| O+AC | 72,84 ± 11,55ns#### | 236,22 ± 30,82ns#### | 113,83 ± 42,25ns# | 39,69 ± 1,28ns | 78,53 ± 3,33ns## |
| N+O | 61,87 ± 23,91ns#### | 181,98 ± 35,55ns#### | 97,66 ± 29,17ns## | 39,86 ± 0,75ns | 78,58 ± 2,92ns## |
| Groups | Necrosis | Congestion | Inflammatory cells | Pyknosis | Portal space |
| N | - | - | - | - | - |
| AC | ++++ | ++++ | +++ | + | + |
| O+AC | - | - | + | - | - |
| N+O | - | - | - | - | - |
| Groups | GSH content | MDA content |
|---|
| Cytosolic Level(µmol/mg proteins) | Mitochondrial level(µmol/mg proteins) | CytosolicLevel (µmol/mg proteins) | Mitochondrial level(µmol/mg proteins) |
|---|
| N | 0,45 ± 0,04 | 0,143 ± 0,0 | 0,566 ± 0,25 | 0,560 ± 0,11 |
| AC | 0,296 ± 0,05* | 0,096 ± 0,005**** | 2,07 ± 0,65**** | 0,860 ± 0,07*** |
| O +AC | 0,619 ± 0,12**#### | 0,137 ± 0,048ns | 1,09 ± 0,45ns## | 0,455 ± 0,06ns#### |
| N+O | 0,584 ± 0,072*#### | 0,162 ± 0,017ns | 0,830 ± 0,27ns### | 0,599 ± 0,04ns## |
| Groups | Catalase activity | GST activity |
|---|
| Cytosolic Level(µmol/mg proteins) | Mitochondrial level(µmol/mg proteins) | CytosolicLevel (IU/mg proteins) |
|---|
| N | 0,11 ± 0,02 | 0,0938 ± 0,04 | 0,101 ± 0,03 |
| AC | 0,026 ± 0,01** | 0,044 ± 0,021* | 0,046 ± 0,01* |
| O +AC | 0,10 ± 0,05ns# | 0,0823 ± 0,01nsns | 0,098 ± 0,029ns# |
| N+O | 0,13 ± 0,02ns### | 0,105 ± 0,01ns## | 0,118 ± 0,02ns## |
| Groups | Optical density at 540nm |
|---|
| N | 1,709 ± 0,09 |
| AC | 1,104 ± 0,28*** |
| O +AC | 1,307 ± 0,27* |
| N+O | 1,481 ± 0,11# |