The present study demonstrated an accelerating effect of harmine on DPSCs differentiation into neurons in two-dimensional and three-dimensional culture systems. The data indicated that alginate hydrogel provided a suitable three-dimensional environment for the DPSCs culture and neural differentiation.
Alginate hydrogel at 1.2% w/v did not affect the viability or proliferation rates of the cells. In the present study, DPSCs were transferred to 1.2% alginate hydrogel and cultured in a neural differentiation medium. Ashton et al. cultured rat neural progenitor cells in 1% alginate hydrogel with poly lactic-co-glycolic acid-aluminum (PLGA-Al) for 7 days and observed a significant increase in the proliferation rate of these cells (
30). In Ashton et al.’s study, the presence of PLGA-Al together with alginate hydrogel may have increased the proliferation rate of rat neural progenitor cells by controlling the degradation of this hydrogel; the type of cells and the concentration of alginate used in the two studies are also different. The behavior of different cells in alginate hydrogels has been studied, and different results have been obtained (
30). These results may be related to the specific cell type or the method used to encapsulate the cells (
31). Since alginate does not interfere with cell function, cells cannot adhere to the alginate network. Cell adhesion is necessary for most cells’ survival, proliferation, differentiation, and gene expression (
32). The non-adhesive nature of the alginate hydrogel promotes cell-cell interaction, which is important for maintaining cell survival and enhancing the functional properties of cells (
33).
In this study, it was found that 5 and 10 µM harmine increased the proliferation of DPSCs. In Title et al.’s study, 4 days of harmine treatment (1, 3.3, 5, and 10 µM) increased pancreatic β-cell proliferation. Although the duration of treatment in the two studies is different, the results are completely consistent (
34). In contrast to our study, one study showed that concentrations of 1, 3, and 10 μM harmine had no effect on osteoblast cell proliferation. The reason for this contradiction is the possibility of a difference in cell type, showing that harmine does not have a general effect (
35).
It was found in this study that 5 and 10 μM harmine increased and accelerated stem cell differentiation. A previous study showed that harmine increased osteoblast differentiation (
35).
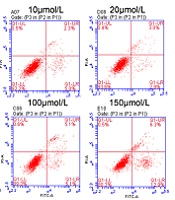
Flow cytometry test indicated that the mean percentage of nestin, MAP-2, and β-tubulin ІІІ positive cells increased in cells treated with harmine compared to the control cells; also, according to the results of the RT-PCR test, the expression levels of all three genes, including
nestin,
MAP-2, and
β-tubulin ІІІ, increased in cells treated with harmine compared to the control cells. This study is the first research to report the accelerating effect of harmine on DPSCs differentiation into neurons. A previous study showed that harmine could increase osteoblast differentiation by inducing bone morphogenetic protein (BMP) expression and activating the BMP and Runx2 signaling pathways (
35).
Encapsulated cells in alginate hydrogel differentiated into neuron-like cells more than cells in two-dimensional cultures, with an increase in the mean percentage of nestin, MAP-2, and β-tubulin ІІІ positive cells compared to the control cells. Also, according to the results of the RT-PCR test, the expression levels of
nestin,
MAP-2, and
β-tubulin ІІІ genes increased in encapsulated cells compared to the control cells. This effect has been reported in previous studies. Banerjee et al. cultured mature neural stem cells isolated from rat hippocampus in alginate at a concentration of 0.25% or 1%. Their experiments showed that when examining the neural differentiation of these cells, the expression level of the β-tubulin Ш marker (neural differentiation marker) was higher in cells cultured in alginate at a concentration of 0.25% (
36). These results were consistent with the results of our recent investigation. Xei et al. also cultured stem cells isolated from rat adipose tissue in peptide hydrogel and neuro-induction medium. Ten days after neural differentiation, the immunocytochemical analysis showed that these cells expressed nestin and bromodeoxyuridine (BrdU) (cell proliferation marker) in the center of the formed cell masses (
37). The results of the present study showed a significant increase in nestin expression in cells encapsulated in alginate hydrogel after 14 days of neural differentiation. Although the type of scaffold used in the two studies and the type of cells are different, it is possible that the hydrophilic state created by both scaffolds contributes to similar results. Read et al. showed that the rat glioma BT4C cell line encapsulated in 1.5% alginate hydrogel, cultured in vivo proliferated, and survived 4 months after culture (
9). In contrast, in more recent experiments, DPSCs encapsulated in 1.2% alginate hydrogel and cultured in vitro showed no proliferation. The difference in cell type and concentration of alginate used, and also the in vivo conditions in Read et al.’s study, may explain the different results in the two studies. Purcell et al. cultured mouse cortical neural stem cells in 1% alginate and observed increased survival and proliferation of these cells from day 7 to day 21, in addition to larger proliferating cell masses expressing nestin (
38). In contrast to these results, the proliferation of DPSCs encapsulated in alginate hydrogel did not change in the present study. In our study, the expression of nestin increased. Likely, the different cell types and concentrations of alginate used in the two studies led to different results. Matyash et al. cultured human and rat neural stem cells on a 1% or 0.2% alginate hydrogel neural medium. These cells had longer neurites than the control group. Moreover, 60 - 70% of the cells were positive for MAP-2 or β-tubulin Ш markers (
39). The results of this research showed that DPSCs encapsulated in 1.2% alginate hydrogel were positive for nestin, β-tubulin ІІІ, and MAP-2.








-2,5-diphenyl-2H-tetrazolium bromide assay. Data are expressed as a percentage of control cells as means ± standard deviation. **; P < 0.01, and ***; P < 0.01 compared to control.")
 nestin, (B) microtubule-associated protein, and (C) β-tubulin III proteins were determined by flow cytometry assay.")
 nestin, (B) MAP-2, and (C) β-tubulin III were determined by flow cytometry assay.")

 <i>nestin</i>, (B) microtubule-associated protein, and (C) <i>β-tubulin</i> III, were determined by real-time polymerase chain reaction assay. Data are expressed as a percentage of control cells as mean ± standard deviation ***; P < 0.01 compared to control.")