Advanced cervical cancer (ACC) is regarded as the second most prevalent malignancy in women besides being the most common female-related cancer in dozens of developing countries (
1). ACC patients are often treated with chemotherapy and radiotherapy followed by brachytherapy. Although chemotherapy and radiotherapy are invariably the main treatments for this type of tumor, the prognosis remains extremely poor (
2). Radiotherapy is reported to be the cause of increased morbidity in ACC cases. Thus, this treatment is given only when the para-aortic nodal disease is confirmed (
3). Hence, a precise evaluation of the extent of the tumor is substantial in order to design the most beneficial treatment. The majority of ACC cases contain human papillomavirus (HPV) infection, but viral infection was not detectable in a small percentage of the examined patients (
4), reflecting the involvement of genetic variations as major risk factors for this disease (
5) beside considering the sensitivity of ACC cells to genotoxic agents (
6). Among ACC cell lines, Hela is the most widely used model cell line and no genomic reference for this cell line has been released so far. The analysis of the genomic content of Hela cells has uncovered a remarkably high level of aneuploidy. Moreover, gene expression patterns related to various cellular pathways (i.e., DNA repair pathways) has been reported to be altered in these cells (
7). The cell death happens following the occurrence of DNA damage (
8). Many DNA lesions are found to trigger the cell death including DNA double-strand breaks (DSBs) (
9). The repair of DNA lesions is essential in preventing the cell death. Drugs that suppress DNA repair responses in cells harboring mutations in DNA repair pathway genes could be efficacious in monotherapy or in combination with other chemotherapeutic agents with the possible advantage of fewer side effects (
10). DSBs are normally repaired by the homologous recombination (HR) pathway that involves RAD51 that requires breast cancer susceptibility proteins (BRCA1) and BRCA2 and poly (ADP-ribose) polymerase 1 (PARP1) (
11). Studies revealed that the deficiencies of other HR-related proteins including CHK2, RAD superfamily, RPA1, ATR, ATM, and CHK1 sensitize the cells to PARP inhibition.
PARP1 has a critical role in more than one DNA repair processes, and small molecule selective inhibitors of PARP (PARPi) have been developed as chemotherapy sensitizers in cancer progression research (
12). Olaparib (AZD2281, Lynparza) was the first PARPi approved by the food and drug administration (FDA) for the treatment of patients with deleterious germline BRCA-mutated advanced ovarian cancer (
13). The growth-inhibitory efficacy of this PARPi was further investigated in other types of cancer cells as monotherapy or in combination with classic platinum anticancer drugs, which resulted in promising but limited outcomes due to the issues of drug resistance (
14). In contrast, significantly lower levels of drug resistance were observed in treating HR-deficient breast cancer cells with AZD2461. In addition, anti-proliferative and apoptosis-inducing effects of AZD2461, as a newly developed structural analog for olaparib, have been recently discovered (
15,
16). On the other hand, histone deacetylase inhibitors (HDACi) have been well established to exert anti-tumor effects on different cancerous human cells in vitro and in vivo (
17). Valproic acid (VPA) is a conventional anti-seizure drug that recently has been demonstrated to have anticancer activities mediated by the selective inhibition of cellular histone deacetylase 1 (HDAC1) (
18). VPA was reported to be able to suppress the cell proliferation as a single agent or in combination with other cell growth inhibitors, specifically in Hela cells (
19,
20).
Combining conventional chemotherapeutic drugs with selective inhibitors of DNA repair pathway has recently gained much attention in the field of selective cancer treatment (
21). While mutations in HR-related genes are associated with much higher risk of breast and ovarian cancers (
22), these genetic variations are not frequent in other cancer types including ACC. The present experiment aimed to evaluate cell viability reducing and DNA repair potential of combined AZD2461 and VPA in Hela cells.








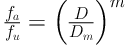
 and in combination (C) depicted by GraphPad Software. Dose-response plot (D), combination index plot (E), isobologram plot (F), median effect plot (G) and polygonogram (H) for co-treatment with AZD2461 and VPA are depicted by Compusyn software. Different effective doses (EDs) of VPA and AZD2461 combination indicate a mild antagonism (CI > 1.1) (G) as Hela cells were treated with various combinations of both inhibitors.")

