
Introduction
Experimental
Results
Discussion
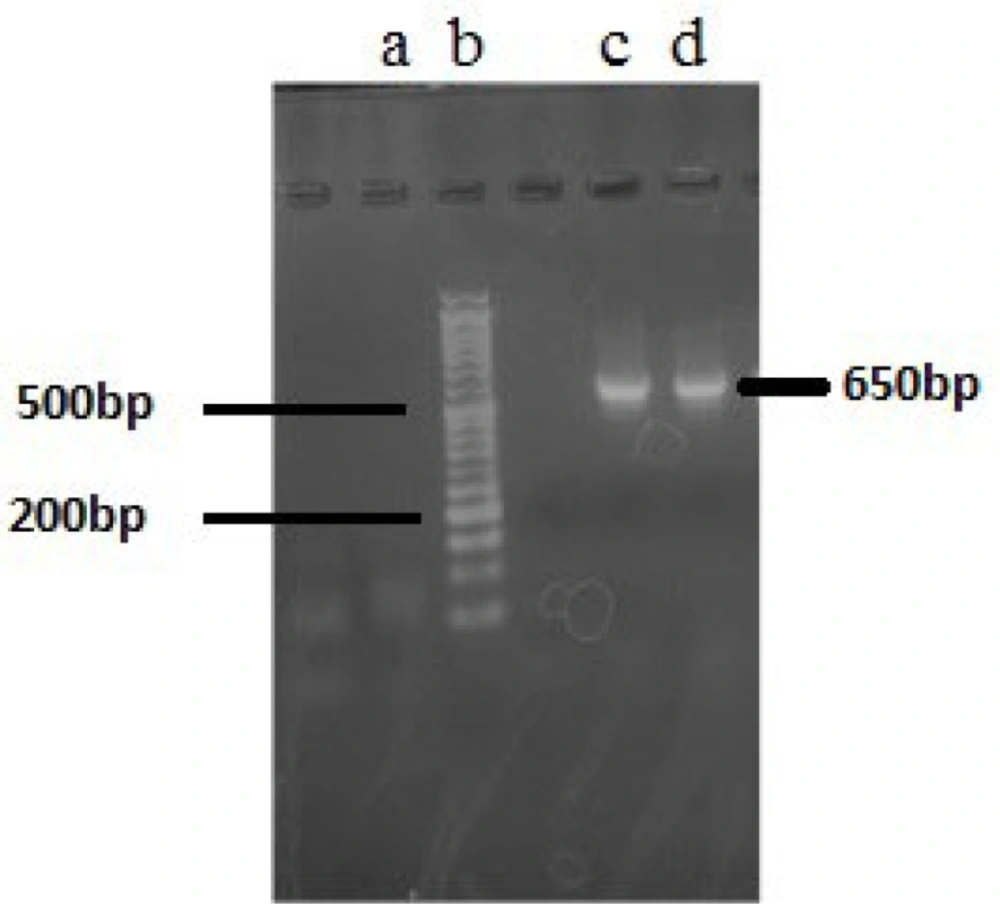
Agarose gel electrophoresis (2%) of 16S rRNA gene PCR products. (a) blank; (b) 1.5 kb DNA ladder; (c) PCR products of F5; (d) PCR product of F14
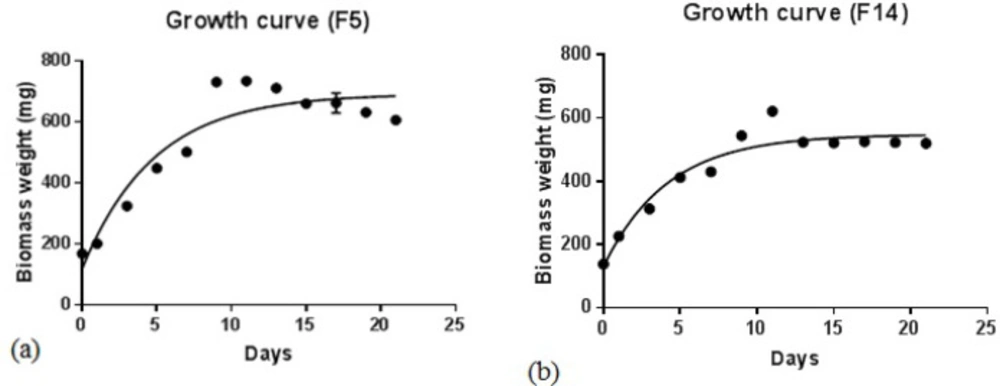
Growth curves of Fischerella species, F5 (a) and F14 (b), in BG-11 culture medium at 25 ºC under 16:8 h light /dark condition
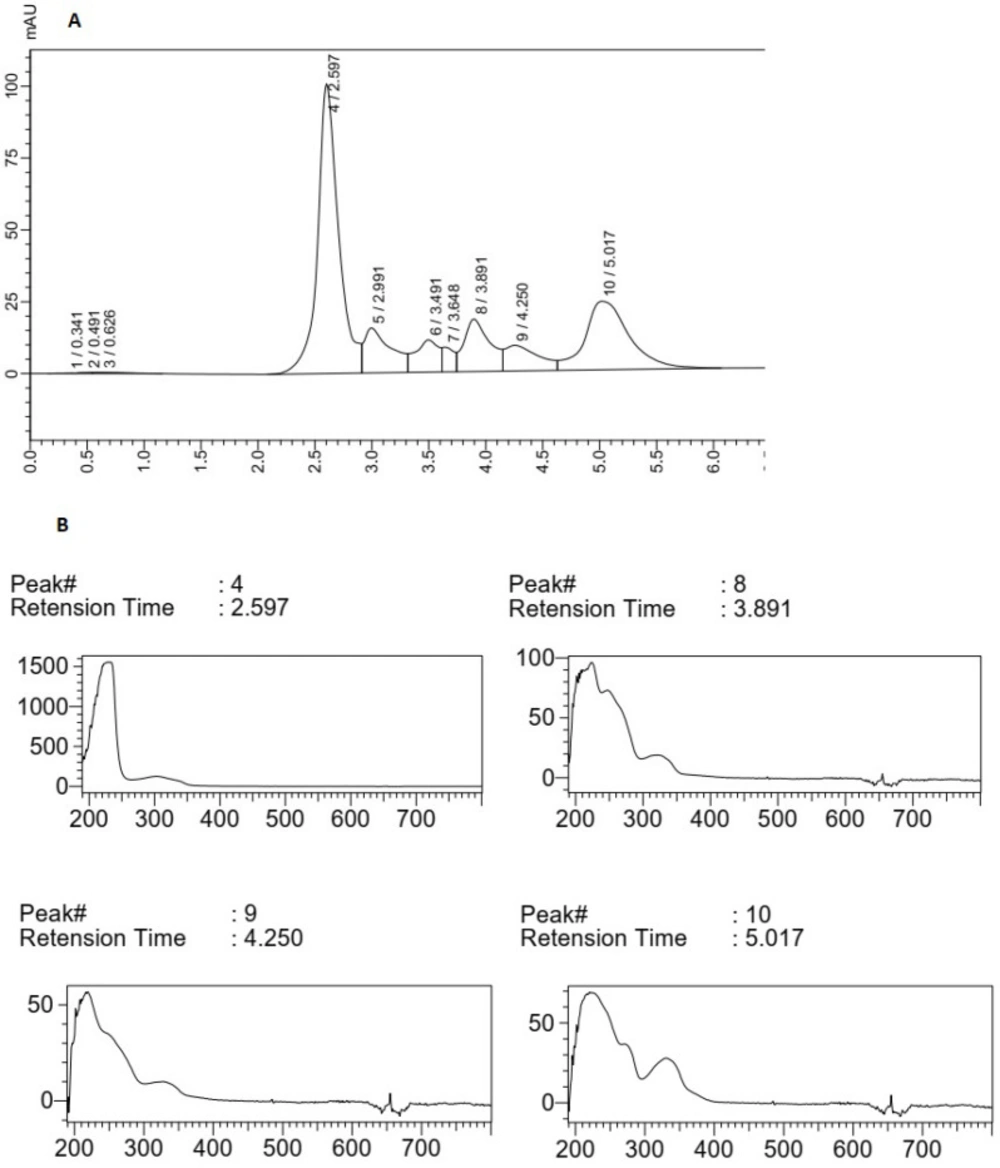
(A) HPLC chromatogram of F5 extract, (B) Peak (4), peaks (8), (9) and (10) correlated with main compounds and the compounds having UV absorption at 300-400 nm
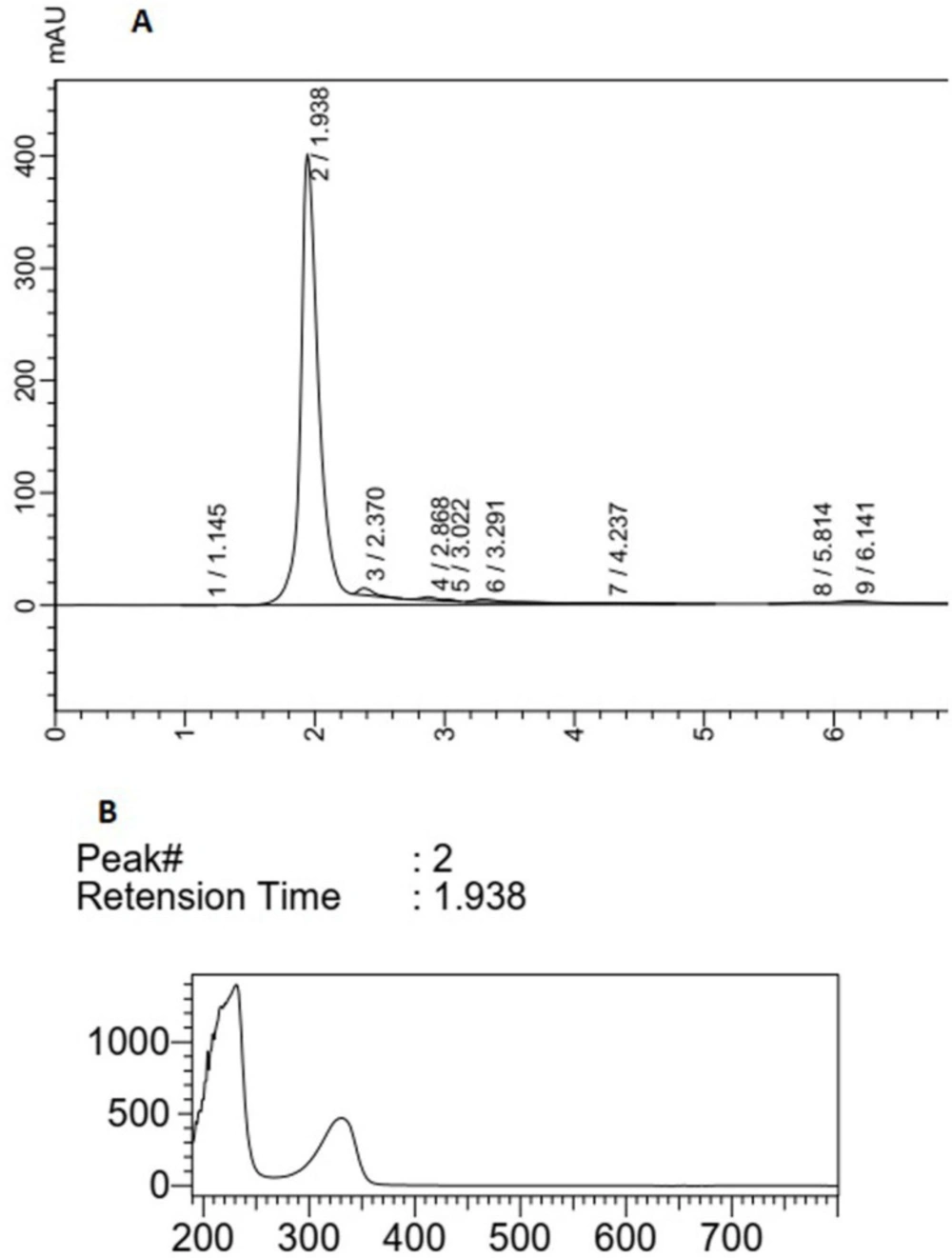
(A) HPLC analysis of F14 extract, (B) peak (2) correlated with compounds having UV absorption at 300-400 nm
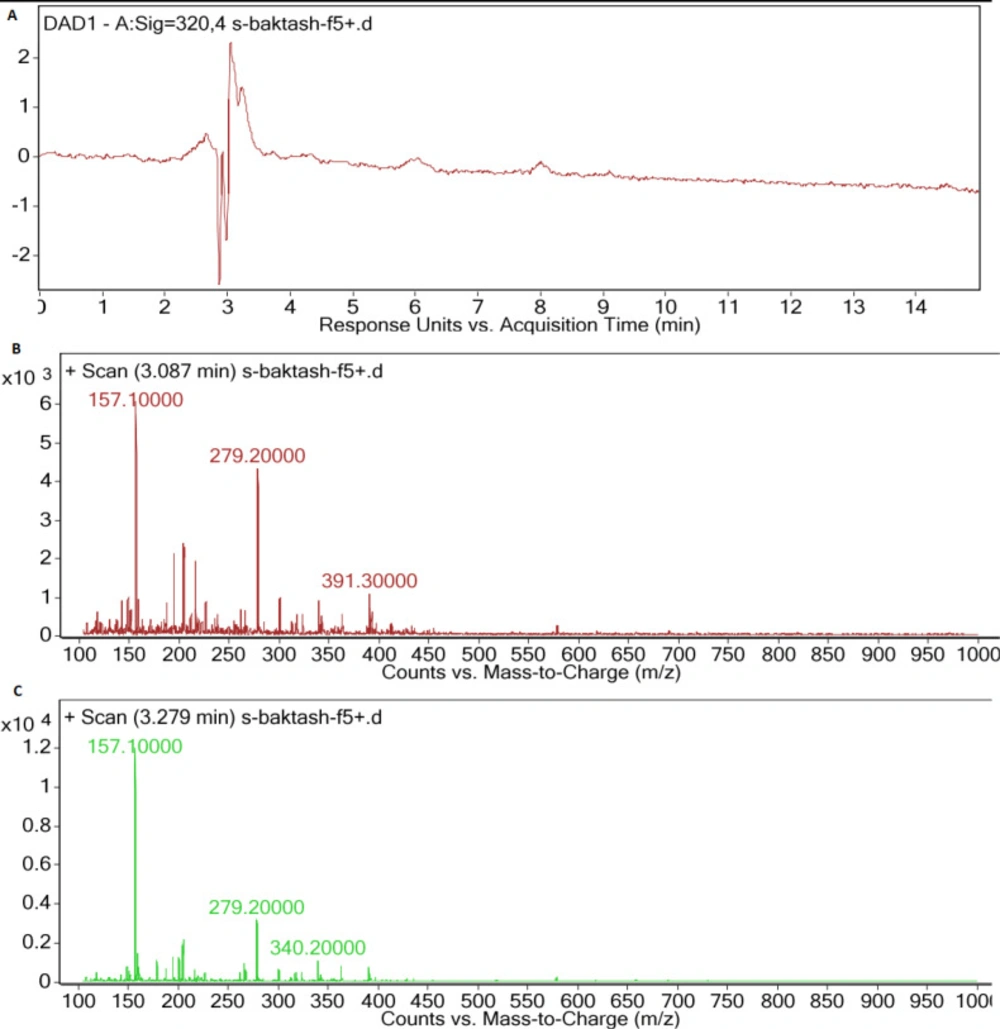
(A) LC-DAD spectra of F5 extract, (B) MS spectrum of main peak (Rt: 3.087 min), (C) MS spectrum of second peak (Rt: 3.279 min), with UV absorption at 300-400 nm
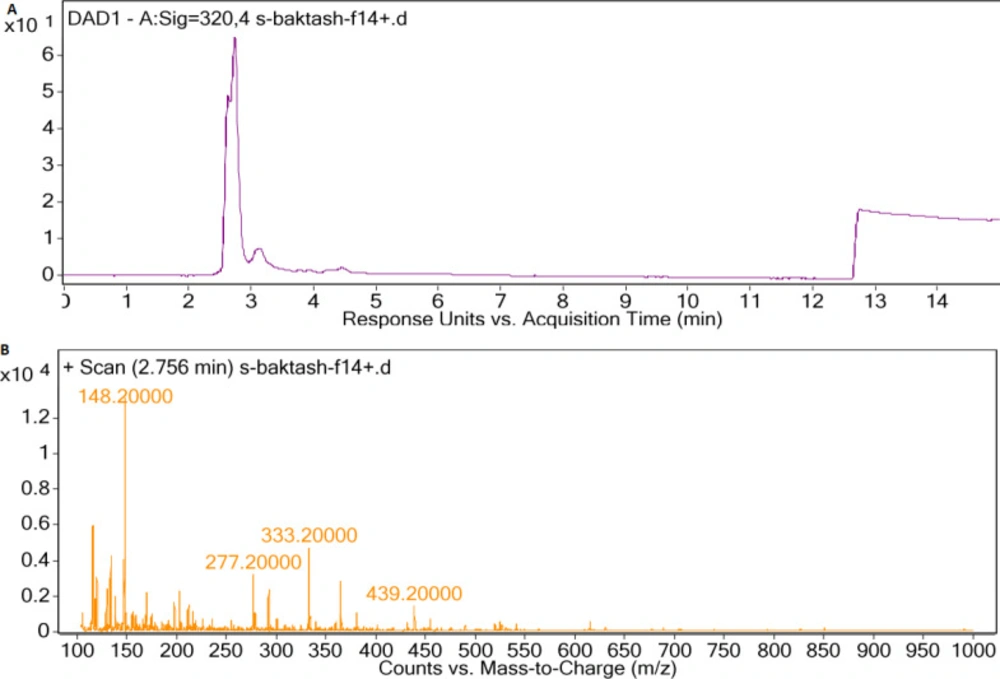
(A) LC-DAD spectra of F14 extract, (B) MS spectrum of main peak with UV absorption at 300-400 nm
| Amplicon | Primer | Sequences | Tm | Site | Ref. |
|---|---|---|---|---|---|
| 630 | CYA106F | CGGACGGGTGAGTAACGCGTG | 65.8 | 106–127 | (16) |
| CYAN738R | GCTAGGACTACWGGGGTAT | 57.5 | 738–765 | ||
| 1260 | CYAN738F | ATACCCCWGTAGTCCTAGC | 57.5 | 738–765 | (17) |
| CYAN1281R | GCAATTACTAGCGATTCCTCC | 57.8 | 1281–1302 |
| Sample | Description | Max Score | Query Cover | Per. Ident | Accession |
|---|---|---|---|---|---|
| F5 | Fischerella sp. Sara 1 16S ribosomal RNA gene, partial sequence | 928 | 94% | 99.61% | KY618863.1 |
| Fischerella sp. MGCY374 16S ribosomal RNA gene, partial sequence | 928 | 94% | 99.61% | KY056817.1 | |
| F14 | Fischerella sp. ATCC 43239 16S ribosomal RNA gene, partial sequence | 627 | 95% | 86.52% | KJ768872.1 |
| Fischerella sp. IR-291 16S ribosomal RNA gene, partial sequence | 627 | 95% | 86.52% | KY011910.1 |
