The term “Apoptosis” derives from the Greek language and means trees shedding their leaves, the leaves “falling off” from trees, in autumn. Apoptosis is crucial for physiological processes, embryonic development, tissue homeostasis, and pathological diseases, such as cancer. Normally, it is triggered by two different molecular pathways comprising the death receptor (extrinsic) and mitochondrial (intrinsic) pathways, both of which converge on the activation of the cascade of caspases (
27). HDACIs induce apoptosis through the activation of both pathways, intrinsic and extrinsic. They activate the extrinsic pathway via the upregulation of death receptors expression, reduction in c-FLIP, and upregulation of ligands such as TRAIL. Further, they activate the intrinsic pathway through the upregulation of several Bcl-2 family genes such as Bid, Bim, and Bmf (
28). Similar to HDACIs, DNA demethylating drugs cause apoptosis via both extrinsic and intrinsic pathways (
29).
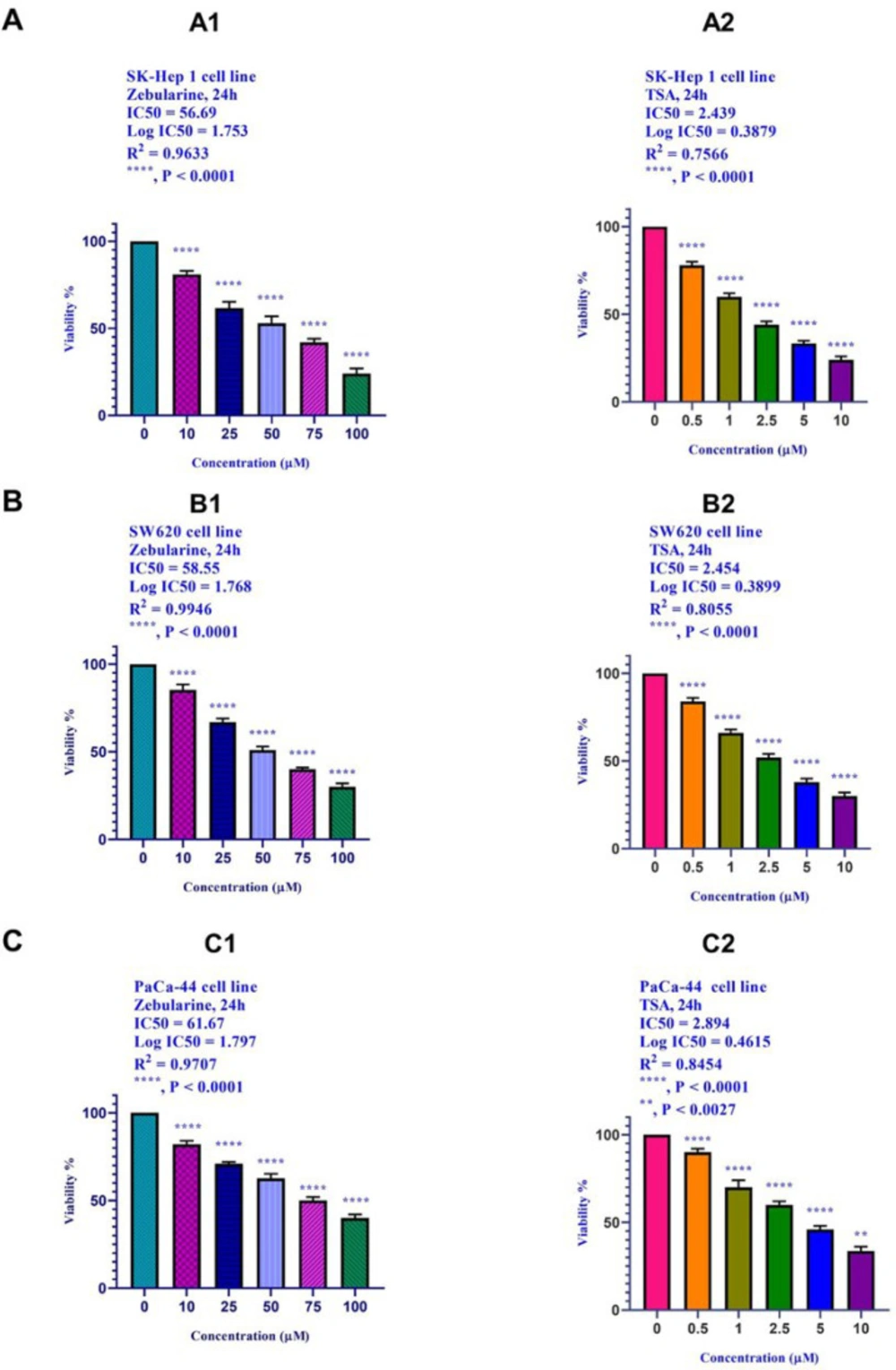
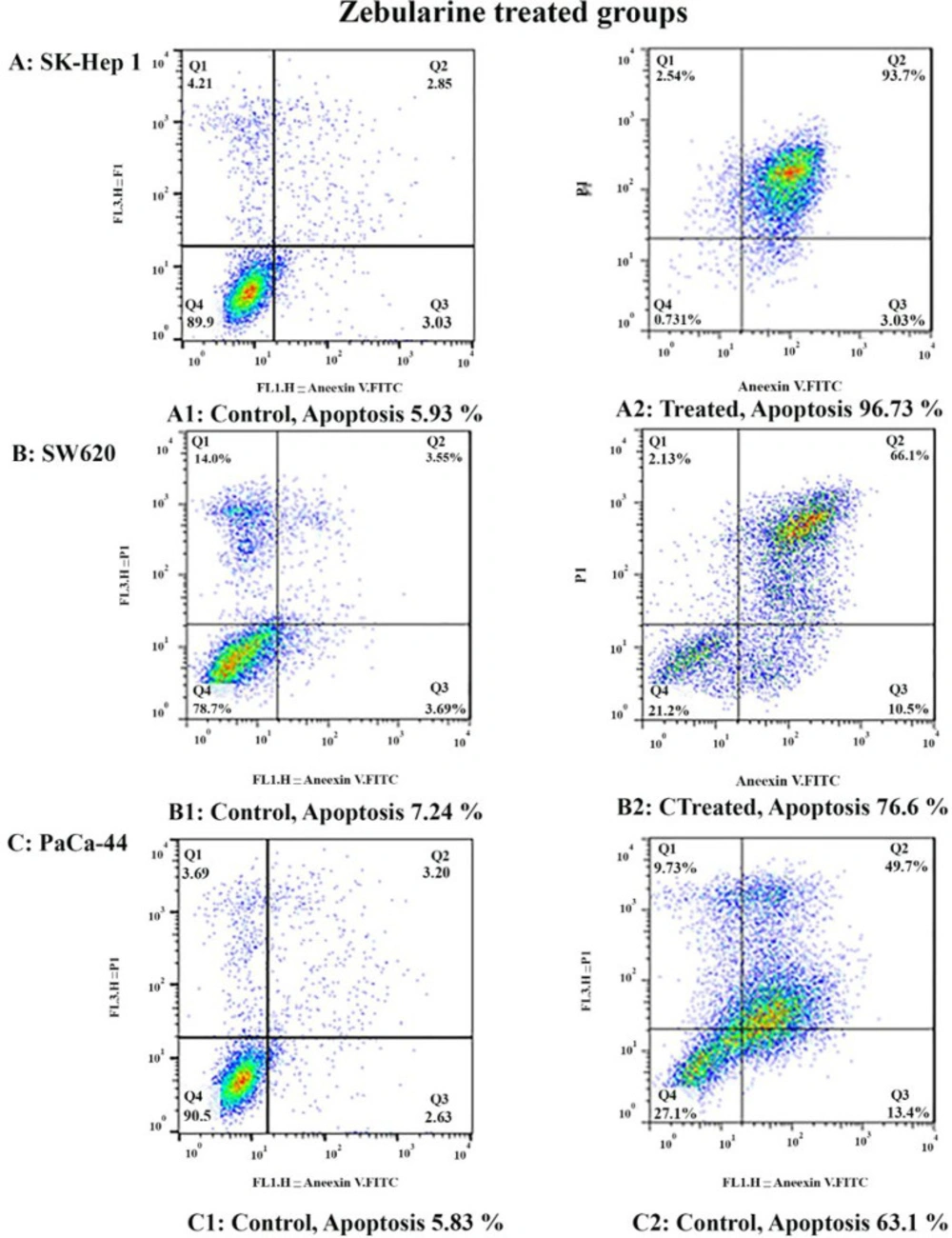
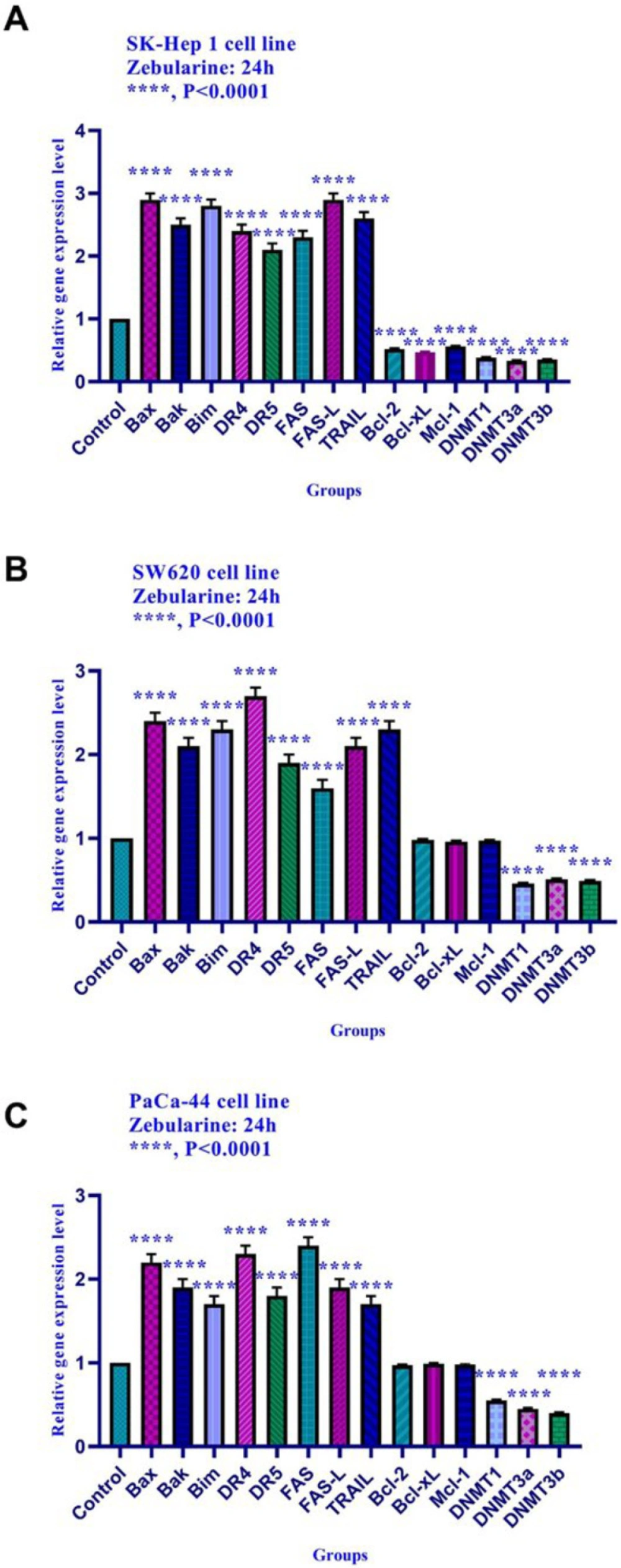
The results of current studies indicated that DNA methyltransferase inhibitor zebularine can induce apoptosis in hepatocellular carcinoma SK-Hep 1, human colorectal cancer SW620, and human PaCa-44 pancreatic cancer cell lines. It up-regulated the expression of the genes of the extrinsic apoptotic pathway and pro-apoptotic genes (as mentioned in the result part) and down-regulated the expression of the anti-apoptotic genes,
DNMTs (1, 3a, and 3b),
HDACs (1, 2, and 3) in SK-Hep 1 cell line significantly. Additionally, zebularine up-regulated the expression of the genes of the extrinsic apoptotic pathway and pro-apoptotic genes (as mentioned in the result part) and down-regulated the expression of DNMTs (1, 3a, and 3b), HDACs (1, 2, and 3) in SW620, and PaCa-44 cell lines significantly. We did not observe significant changes in the expression level of
Bcl-2, Mcl-1, and Bcl-xL in SW620, and PaCa-44 cell lines treated with zebularine. Similarly, in-vitro studies have demonstrated that DNA demethylating agents zebularine and decitabine activate the mitochondrial apoptotic pathway in T cells (
30). Similar to our findings, it has been reported that zebularine treatment induces cell-cycle arrest, and apoptosis in the HCC HepG2 cell line through the intrinsic pathway (
31). It has been reported that DNA methyltransferase inhibitor 5-Aza-CdR down-regulated MCL-1 leads to apoptotic induction in AML cells (
32). In recent studies, it was demonstrated that zebularine and 5-aza-dC down-regulate
DNMT-1, DNMT-3a, and DNMT-3b in ovarian cancer cell lines (
33). Several works have demonstrated that 5-Aza-CdR treatment decrease
DNMT1 and DNMT3a mRNA expression resulting in a down-regulation of Bcl2 protein in colon cancer cell line HCT-116 (
34). Experimental studies have shown that compounds with DNMTI activity such as 5-Aza-CdR generally induce tumor cell apoptosis by DNA demethylation in human cell lines A549, HepG2, Hep3B, A-498, and HCT-116 (
35).
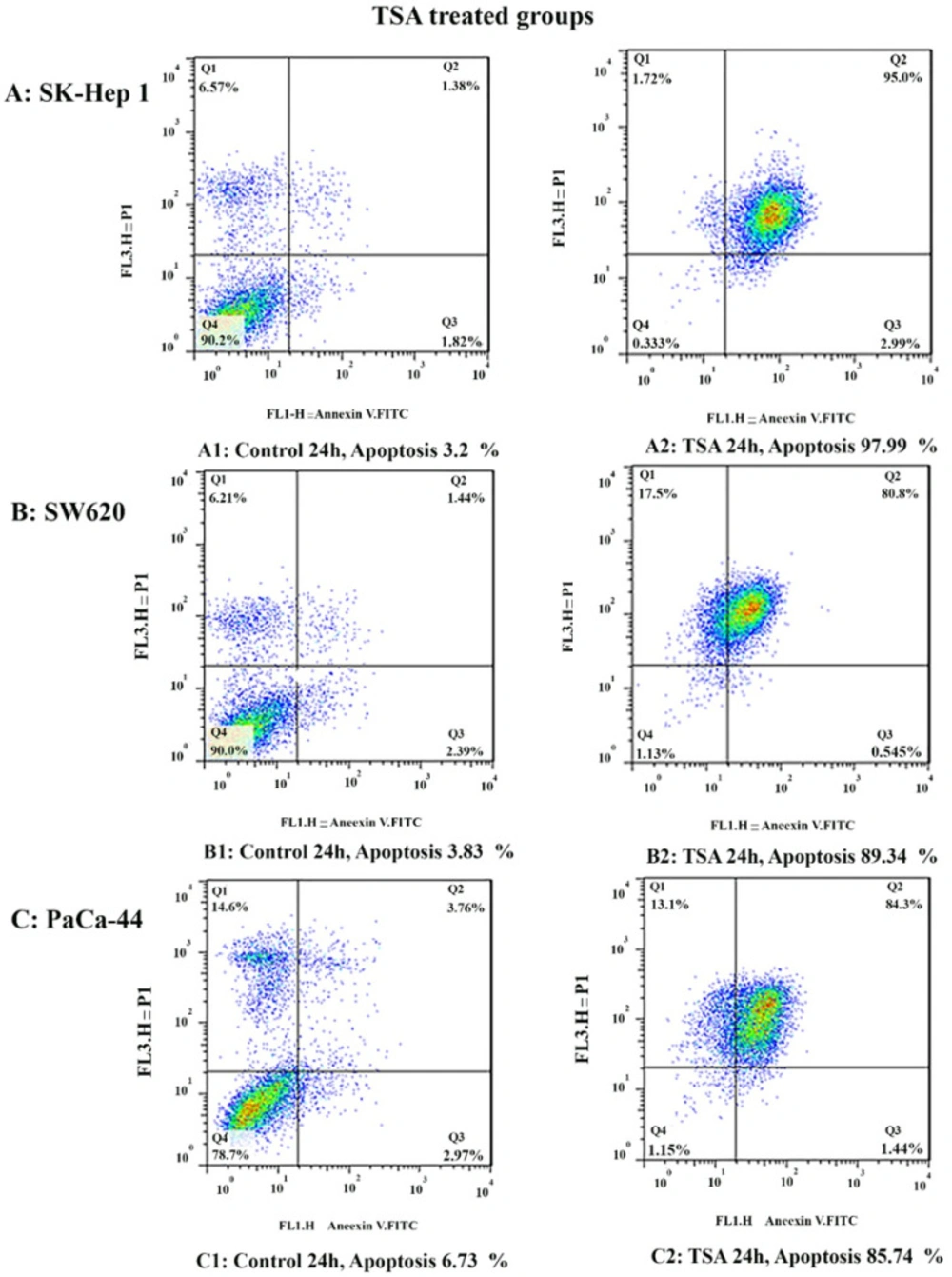
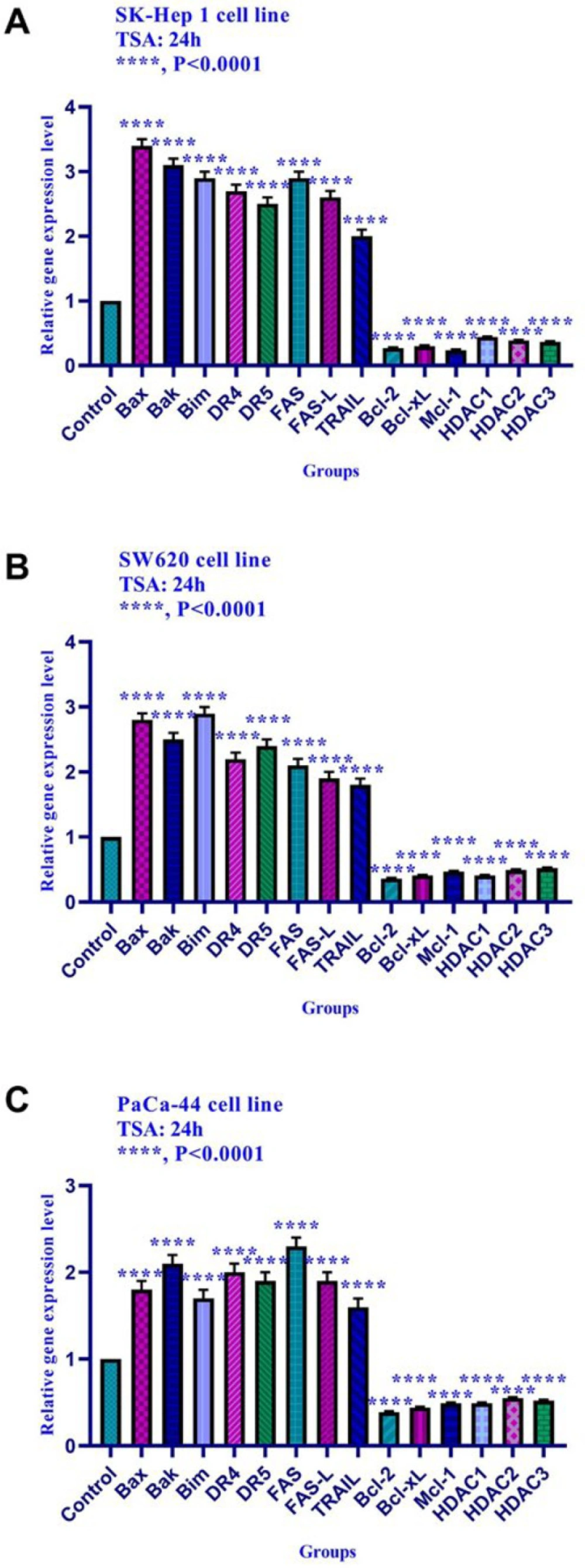
In the present work, we reported that histone deacetylase inhibitor TSA can induce apoptosis through both mitochondrial/intrinsic and cytoplasmic/extrinsic apoptotic pathways, in hepatocellular carcinoma SK-Hep 1, human colorectal cancer SW620, and human PaCa-44 pancreatic cancer cell lines through the activation of both extrinsic and intrinsic pathways, it up-regulated the expression of
Bax, Bak, Bim, DR4, DR5, FAS, FAS-L, and TRAIL genes and down-regulated the expression of
Bcl-2, Mcl-1, Bcl-xL, DNA methyltransferase 1, 3a, and 3b, histone deacetylase 1, 2, and 3 in all three cell lines significantly. Inconsistent with our result, other researchers have indicated that histone deacetylase inhibitor sodium butyrate decreases Bcl-XL protein levels by down-regulating its RNA expression in mesothelioma cell lines. Besides, depsipeptide, as an HDACI, decreases the expression of
Bcl-2, Bcl-XL, and Mcl-1 in multiple myeloma cells (
36). In human lymphatic endothelial cells (LEC), TSA-induced apoptosis by cytochrome c release contributed to activating caspases-3, caspases −7, and caspases −9 and the anti-apoptotic proteins down-regulation accompanied by up-regulation of
p21, p27, and p53 (
37). Resent in-vitro works have shown that TSA increases the ratio between the levels of expression of anti-apoptotic (
BCL-w and BCL-xl) and pro-apoptotic (BIM)genes in pancreatic cancer cell lines CFPAC1, Miapaca2, HPAF, PSN1, Panc1 PC, Paca44, RT45P1, and T3M4 (
38). Similar to our findings, other researchers have shown that HDACIs play their apoptotic roles through the extrinsic apoptotic pathway. It has been indicated that several HDACIs like SAHA, TSA, m-carboxycinnamic acid bishydroxamide (CBHA), LAQ824 (a cinnamic acid hydroxamate), and MS-275 can up-regulate the expression of
TRIAL-R1 and TRIAL-R2 (
39). In pancreatic cancer cell lines Panc1 and MiaPaCa2, treatment with TSA increase the expression of the TRAIL receptor 1 (DR5) (
40). Additionally, HDACIs can induce apoptosis through activation of the death receptor pathway (TRAIL and Fas signaling pathways), the up-regulation of TRAIL, DR5, Fas, Fas-L in leukemic blasts (
41).
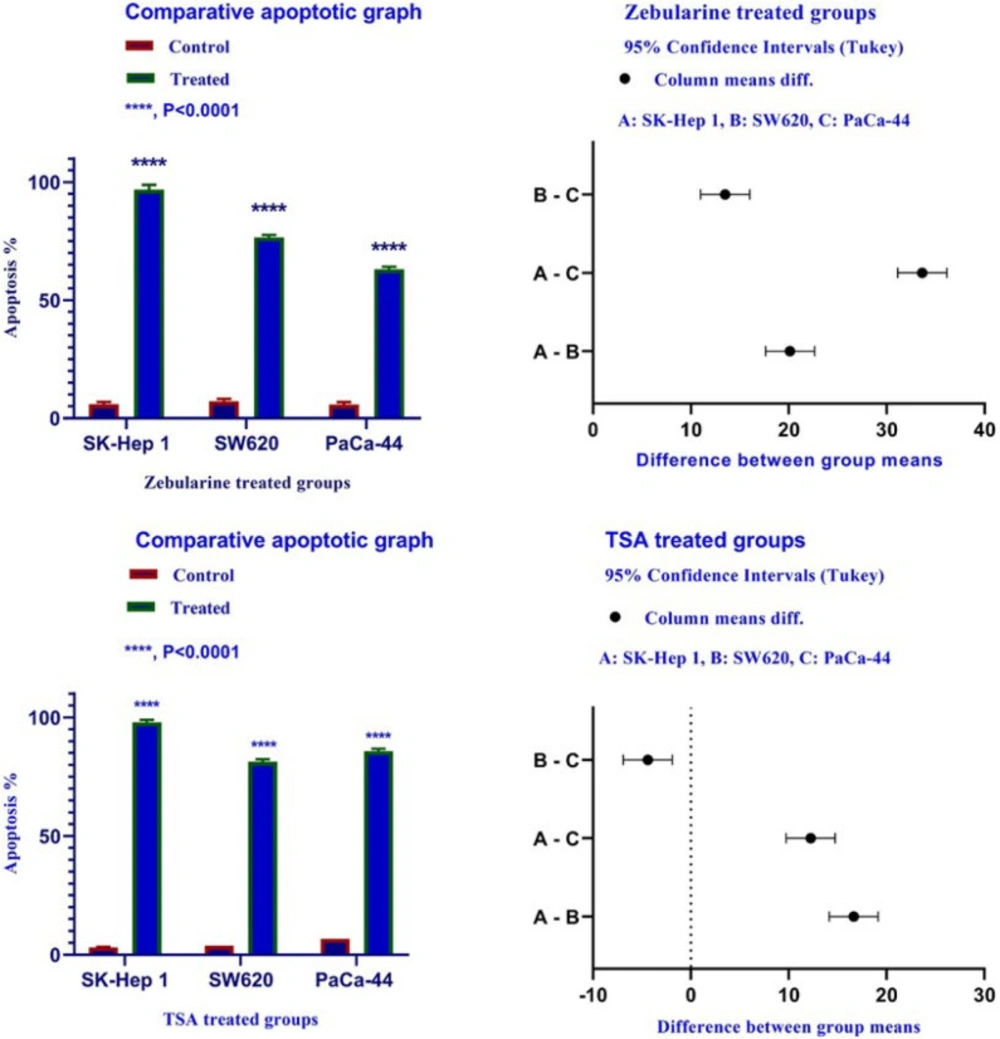
Meanwhile, the extrinsic and intrinsic pathways are not the only molecular mechanisms of zebularine and TSA. The results of current studies indicated that DNA methyltransferase inhibitor zebularine and histone deacetylase inhibitor TSA can induce apoptosis through both mitochondrial/intrinsic and cytoplasmic/extrinsic apoptotic pathways, in hepatocellular carcinoma SK-Hep 1, human colorectal cancer SW620, and human PaCa-44 pancreatic cancer cell lines. Our finding demonstrated that zebularine induced significant apoptosis in all three cell lines. It up-regulated the expression of
Bax, Bak, Bim, DR4, DR5, FAS, FAS-L, and TRAIL genes and down-regulated the expression of
Bcl-2, Mcl-1, Bcl-xL, DNA methyltransferase 1, 3a, and 3b, histone deacetylase inhibitors 1, 2, and 3 in SK-Hep 1 cell line significantly. Additionally, zebularine up-regulated the expression of
Bax, Bak, Bim, DR4, DR5, FAS, FAS-L, and TRAIL genes and down-regulated the expression of
DNA methyltransferase 1, 3a, and 3b in SW620, and PaCa-44 cell lines significantly. It had no significant effect on
Bcl-2, Mcl-1, and Bcl-xL gene expression in SW620, and PaCa-44 cell lines. Similarly, in-vitro studies have demonstrated that DNA demethylating agents zebularine and decitabine activate the mitochondrial apoptotic pathway in T cells (
30). Similar to our findings, it has been reported that zebularine treatment induces cell-cycle arrest, and apoptosis in the HCC HepG2 cell line through the intrinsic pathway (
31). It has been reported that DNA methyltransferase inhibitor 5-Aza-CdR down-regulated MCL-1 leads to apoptotic induction in AML cells (
32). In recent studies, it was demonstrated that zebularine and 5-aza-dC down-regulate
DNMT-1, DNMT-3a, and DNMT-3b in ovarian cancer cell lines (
33). Several works have demonstrated that 5-Aza-CdR treatment decrease
DNMT1 and DNMT3a mRNA expression resulting in a down-regulation of Bcl2 protein in colon cancer cell line HCT-116 (
34). Experimental studies have shown that compounds with DNMTI activity such as 5-Aza-CdR generally induce tumor cell apoptosis by DNA demethylation in human cell lines A549, HepG2, Hep3B, A-498, and HCT-116 (
35). As mentioned in the result section, TSA induced significant apoptosis in all three cell lines, SK-Hep 1, SW620, and PaCa-44. It up-regulated the expression of
Bak, Bax, Bim, DR4, DR5, FAS, FAS-L, and TRAIL genes and down-regulated the expression of
Bcl-2, Bcl-xL, Mcl-1, histone deacetylase inhibitors 1, 2, and 3 in these cell lines significantly. Inconsistent with our result, other researchers have indicated that histone deacetylase inhibitor sodium butyrate decreases Bcl-XL protein levels by down-regulating its RNA expression in mesothelioma cell lines. Besides, depsipeptide, as an HDACI, decreases the expression of
Bcl-2, Bcl-XL, and Mcl-1 in multiple myeloma cells (
36). In human lymphatic endothelial cells (LEC), TSA-induced apoptosis by cytochrome c release contributed to activating caspases-3, caspases −7, and caspases −9 and the anti-apoptotic proteins down-regulation accompanied by up-regulation of
p21, p27, and p53 (
37). Resent in-vitro works have shown that TSA increases the ratio between the levels of expression of anti-apoptotic (
BCL-w and BCL-xl) and pro-apoptotic (BIM)genes in pancreatic cancer cell lines CFPAC1, Miapaca2, HPAF, PSN1, Panc1 PC, Paca44, RT45P1, and T3M4 (
38). Similar to our findings, other researchers have shown that HDACIs play their apoptotic roles through the extrinsic apoptotic pathway. It has been indicated that several HDACIs like SAHA, TSA, m-carboxycinnamic acid bishydroxamide (CBHA), LAQ824 (a cinnamic acid hydroxamate), and MS-275 can up-regulate the expression of TRIAL-R1 and TRIAL-R2 (
39). In pancreatic cancer cell lines Panc1 and MiaPaCa2, treatment with TSA increase the expression of the TRAIL receptor 1 (DR5) (
40). Additionally, HDACIs can induce apoptosis through activation of the death receptor pathway (TRAIL and Fas signaling pathways), the up-regulation of TRAIL, DR5, Fas, Fas-L in leukemic blasts (
41). Meanwhile, the extrinsic and intrinsic pathways are not the only molecular mechanisms of zebularine and TSA. Our previous work indicated that zebularine and TSA can inhibit DNMTs (
DNMT1, DNMT3a, and DNMT3b), Class I HDACs (
HDACs 1, 2, 3), and Class II HDACs (
HDACs 4, 5, 6) and up-regulate CIP/KIP Family (
p21Cip1/Waf1/Sdi1, p27Kip1, and p57Kip2) resulting in cell apoptosis in colon cancer LS 174T, and LS 180 cell lines (
42,
43). Finally, zebularine and TSA can play their apoptotic roles through intrinsic and extrinsic pathways. In some cases, zebularine could not change the gene expression significantly. It may induce a significant change in the expression of the mentioned genes with high concentrations at 48 or 72 h. It may be done with further concentrations or durations. Therefore, the evaluation of this compound with high concentrations and more duration (24 and 48h) is recommended.