In this study, the effects of KGN, ASU and TGF-β3 on the process of chondrogenic differentiation in hADSCs on fibrin scaffold were investigated and compared in two laboratories and animal models. Undoubtedly, articular cartilage defects are one of the most challenging issues in the field of medical science and diseases are considered synovial joint. Due to the intrinsic nature of the cartilage tissue, which is the lack of blood vessels and nerves in the tissue, the repair of this tissue is slow and almost insignificant, apart from a complete lack of healing to activate a subset of inflammatory agents involved in cartilage defects (
21,
22). Today, tissue engineering and regenerative medicine aimed to replace lost organs with evolve biotechnologies that combine biomaterials, growth factors, and stem cells. The use of stem cells in the treatment of various diseases is growing.
Among available stem cell sources, ADSCs are an accessible and abundant resource that can be used in cognitive medicine. In 2002, Zuk
et al. reported that ADSCs differentiate into Cartilage cells under the influence of growth factor β-TGF1 for 14 days was associated with the production of sulfated proteoglycans, which is consistent with our results. Also, after day 14, it was found that the induced cells showed
Coll II genes,
AGG and
Coll X, which is consistent with the current study (
23).
Other important components in tissue engineering are cartilage scaffolds that have similar properties to the extracellular matrix. Fibrin scaffolding is one of the natural biomaterials that can be autologous extracted from the patient’s blood (
18). This scaffold has been widely used in tissue engineering due to its good biodegradability and biocompatibility, lack of toxicity, enabling cell proliferation and migration, as well as high elasticity (
24,
25).
In a 2011 study by Girandon
et al., to compare proliferation and survival, ADSCs in both fibrin and alginate scaffolds demonstrated that proliferation and survival rates were higher in fibrin scaffolds than in alginate scaffolds. Similarly, apoptosis was lower in this scaffold than in alginate scaffolds (
26).
In this study, we used KGN, ASU and TGF-β3 for chondrogenic differentiation. The purpose of these compounds was to find a suitable compound with good chondrogenic differentiation, reasonable price and no harmful side effects, such as causing hypertrophy and reducing stem cell biology.
In the present study, the use of the TGF-β3 growth factor showed that this growth factor could induce chondrogenic differentiation both in-vitro and in an animal model, which confirms the expression of genes involved in chondrogenesis such as
Coll II,
SOX9 and
AGG. The results of
in-vitro expression of these genes and observation of Coll II and X proteins
in-vitro and in the animal model showed that they were significantly effective against ASU, but this increase was not significant for KGN. Expression of the
Coll X gene that indicates hypertrophy in differentiated cells in cells treated with chondrogenic media The differentiated TGF-β3 containing cells were significantly increased both in-vitro and in the animal model compared to the other groups, which means that the TGF-β3 growth factor despite its strong influence on chondrocytes stem cells differentiation. It can lead the cells to hypertrophy and possibly apoptosis, which is one of the disadvantages of using this growth factor. We have found in previous research that TGF-β3 increases cell hypertrophy and promotes cartilage to become bony (
18). In the present study, the increase in Coll X protein was increased in differentiated cells in the medium containing TGF-β3 in-vitro and in the animal model compared to ASU and KGN groups, which seems to increase the use of TGF-β3 growth factor alone.
In the present study, ASU was also used as an inducer of chondrogenesis. ASU is a complex combination of many compounds, including fat-soluble vitamins, sterols, triterpenols, and possibly fatty acids (
27). Past
in-vitro and
in-vivo studies have shown that ASU has a positive effect on osteoarthritis (
28) and inhibits cartilage fracture, and stimulates synthesis by inhibiting a number of molecules and pathways involved in osteoarthritis. It becomes collagen and AGG (
29). It can also prevent the progression of osteoarthritis by inhibiting inflammatory cytokines such as IL-1, IL-6, IL-8, TNF, and PGE2 by modulating NF-kappaB (
30).
Our results showed that after 14 days of ASU
in-vitro, increased expression of genes involved in chondrogenesis, but this increase was not significant compared to other groups group. Previous research has shown that after 21 days, the expression of genes such as
Coll II and
AGG have increased significantly, which is likely due to prolonged use of this factor (
7). Decreased expression of
Coll X can be attributed to the strengths of this inducible factor. This was observed both
in-vitro and in animal models. Stem cell viability was also higher in the ASU group than in the TGF-β3 and KGN groups, although the molecular process of this event is unclear. It appears that the inhibition of inflammatory factors reported by other researchers prevented cell death and increased cell viability has been fundamental. In 2006, YE Henrotin
et al. reported that ASU inhibits stem cell differentiation into bone cells, which is exerted through the process of inhibiting inflammatory factors such as matrix metalloproteinase (MMP) and 2-cyclooxygenase and in turn expressing type
Coll I. X decreases, which is consistent with current research (
31).
Nowadays, the small-molecule drug KGN has been suggested as an inducible factor in cartilage tissue engineering, which requires more extensive research. Johnson
et al. Demonstrated that KGN by binding to the core-binding factor (CBF) -β inhibits the release of this factor from type A and its binding to runt-related transcription factors 1 (RUNX1) and ultimately to the expression of genes involved in chondrogenesis such as
SOX9,
Coll II, and
AGG. It is expressed in rat brain stem cells (
10). Scientists’ research suggests that the effect of KGN is on the chondrogenesis process by activating RUNX1. RUNX1 is a protein family of Runt-related transcription factors that RUNX2 and RUNX3 are other members of this family. Their activation initiates DNA replication (
32,
33). The results of the present study showed that the use of KGN induces the expression of
Coll II,
AGG and
SOX9 genes both
in-vitro and
in-vivo. Past research has shown that KGN decreases
Coll X gene expression by altering the balance from pre-hypertrophy to pre-differentiation and replication (34). In 2014, J.Zhang stated that KGN inhibits the effects of inflammatory factors such as interleukin 1 (IL1) and Tumor necrosis factor β (TNFβ) on chondrocytes in culture and reduces the degenerative process in these cells (
16).
SOX9 gene expression was significantly increased in the KGN group compared to the ASU group. However, in the case of
AGG and
Coll II and
Coll X genes expression, this increase was not significant.
In this study, after 14 days of
in-vitro chondrogenic differentiation of these cells and the fibrin scaffold in which the cells were differentiated, they were transferred to the skin of male rats, and after 14 days of sampling, histological and immunohistochemical samples were taken the experiment found that the cells had more clear cartilage tissue characteristics than laboratory conditions. In 2008, S. Munirah stated that the amount of glycosaminoglycan in scaffolds containing rabbit chondrocytes was significantly increased after implantation under the skin of mice (
35). In 2001, SasanoY
et al. the expression of
Coll I in differentiated cells in the animal model was lower than in the laboratory model (
36). Comparison of the results of specific staining
in-vitro and animal models also showed that the differentiation of cells under the skin increased the amount of cartilage matrix constituents such as proteoglycans in the implanted scaffolds and the accumulation rate. Coll X, indicating the onset and occurrence of hypertrophy in differentiated cells, was significantly decreased in some groups, particularly ASU.
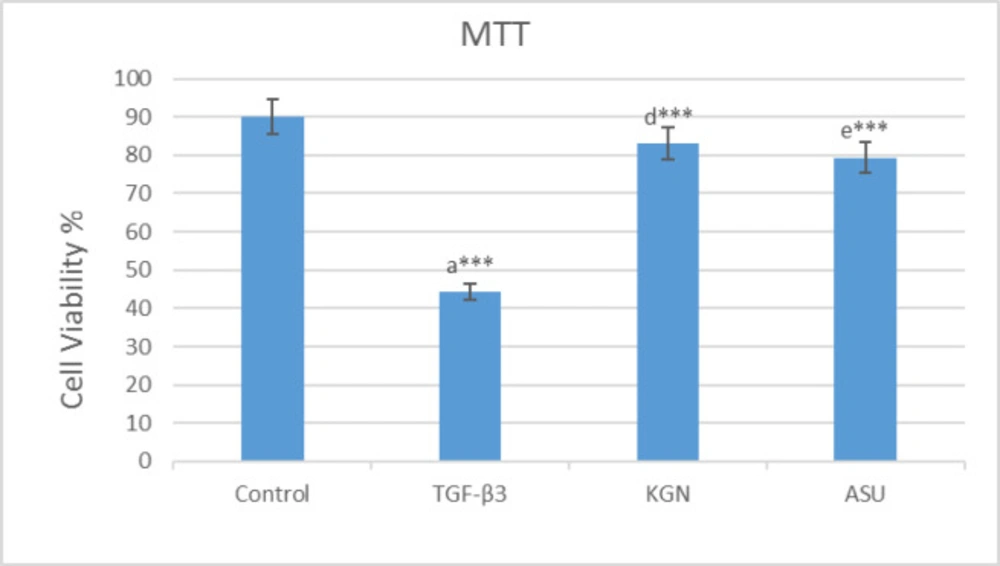
MTT assay results in 14 days after the culture of hADSCs in chondrogenic medium supplemented with TGF-β3, KGN and ASU fibrin scaffold. *P < 0.05, **P < 0.01, ***P < 0.001. a: Difference between control and TGF-β3. b: Difference between control and KGN. c: Difference between control and ASU. d: Difference between TGF-β3 and KGN. e: Difference between TGF-β3 and ASU. f: Difference between KGN and ASU
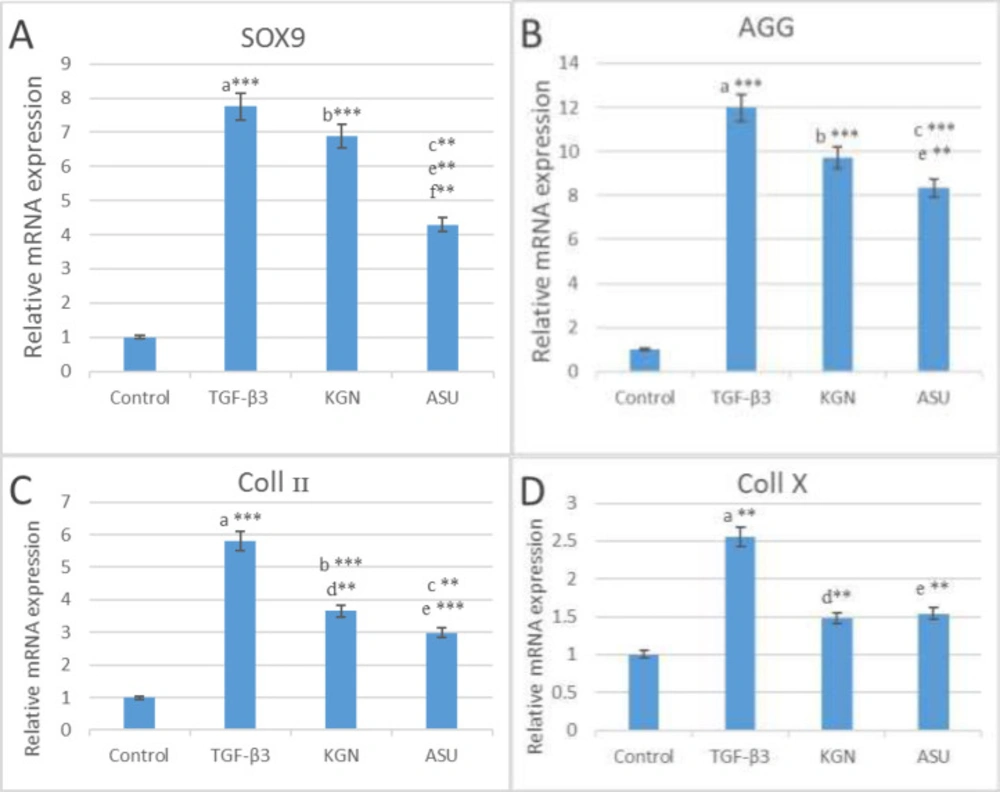
The results of (A) SOX9, (B) AGG, (C) Coll II and (D) Coll X genes expression in control, TGF β3, KGN and ASU groups 14 days after the culture of hADSCs. Data are presented as mean ± SD. Error bars represent the standard deviation of the mean. *P < 0.05, **P < 0.01, ***P < 0.001. RQ (Relative quantification) indicates the relative level of gene expression. aDifference between control and TGF-β3. bDifference between control and KGN. cDifference between control and ASU. dDifference between TGF-β3 and KGN. eDifference between TGF-β3 and ASU. fDifference between KGN and ASU
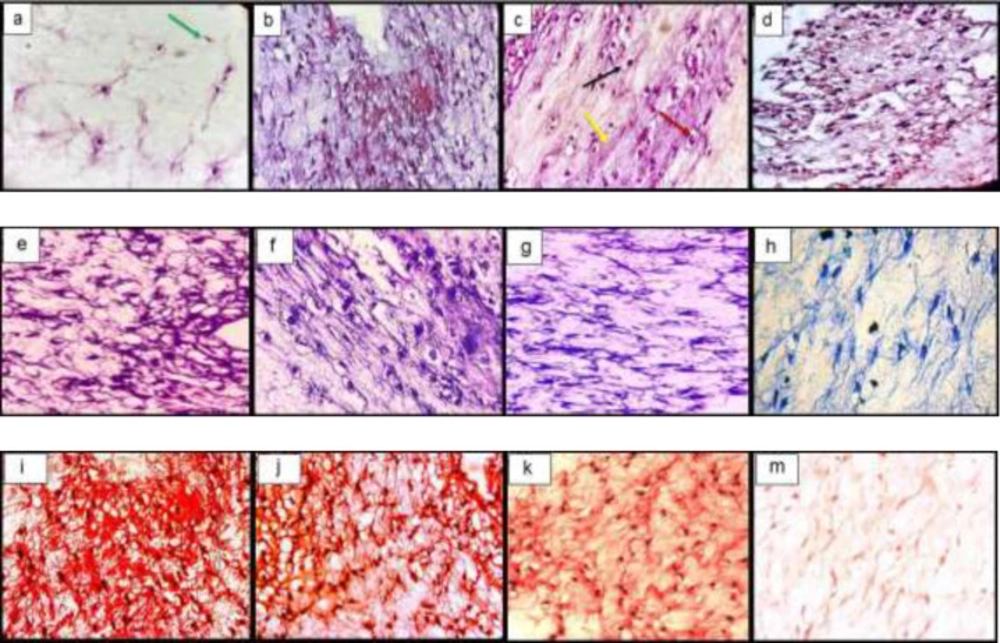
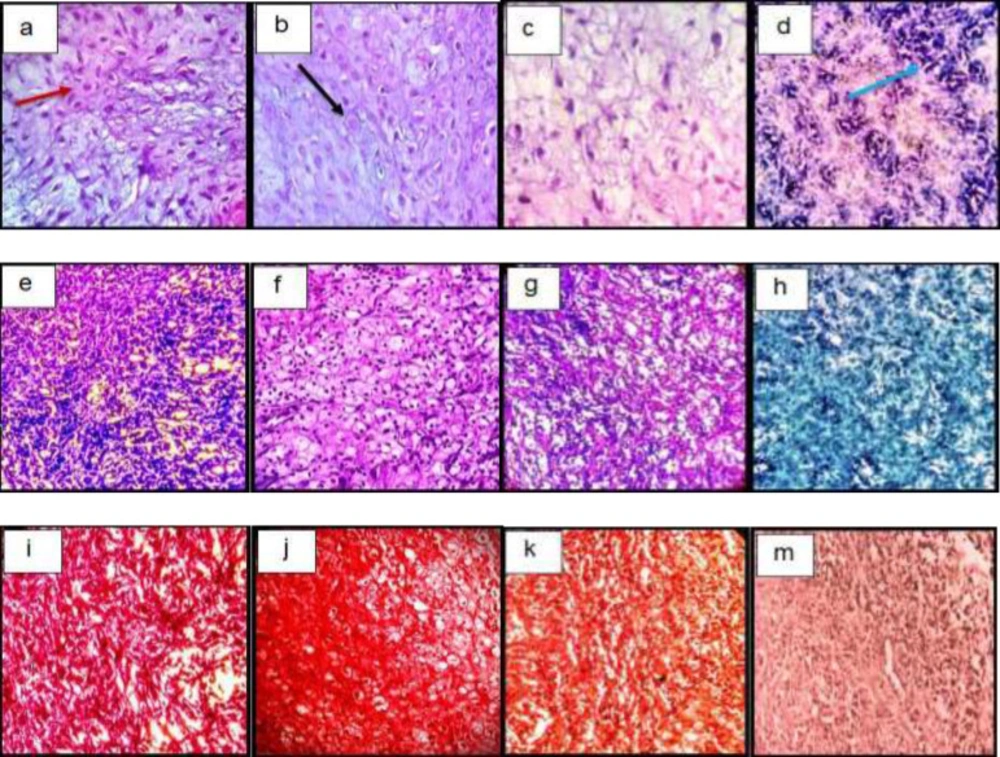
Histological sections of neo-cartilage formed by TGF-β3, KGN and ASU in fibrin scaffold cultures after 14 days as determined by H&E (a: TGF-β3, b: KGN c: ASU and d: Control), Toluidine blue (e: TGF-β3, f: KGN g: ASU and h: Control), Safranin O (i: TGF-β3, j: KGN k: ASU and m: Control) in vitro culture, magnification ×40. Green arrow: Stem cell with an elongated and fusiform nucleus. Red arrow: An isogonic group within Lacuna. Black Arrow: Chondrocytes-like cells inside the lacunae. Yellow arrow: Extracellular matrix
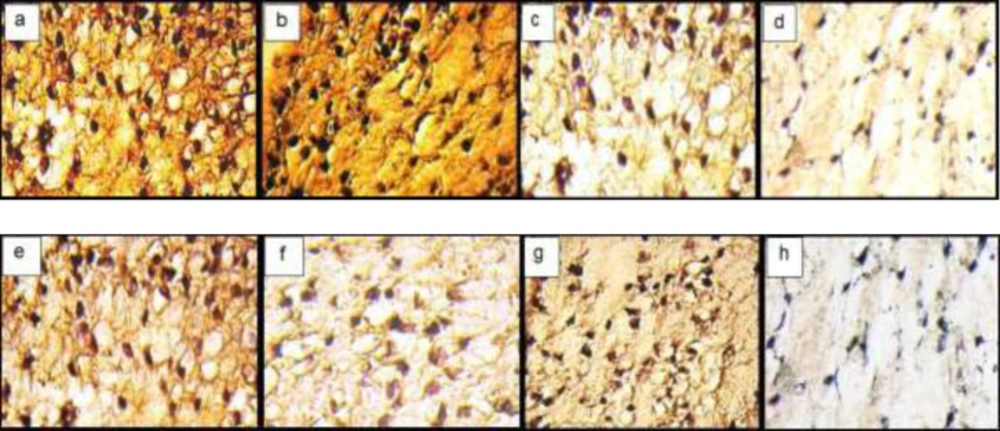
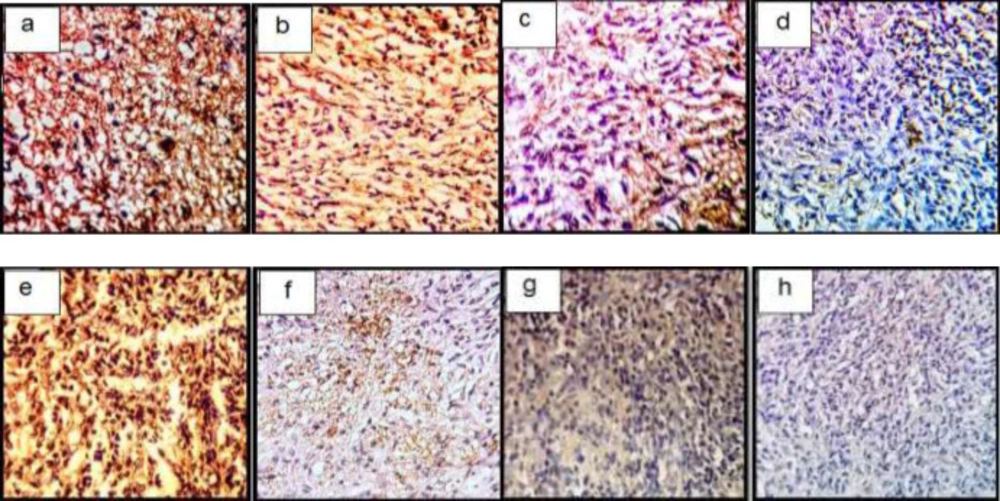
Immunohistochemical sections of neo-cartilage formed by TGF-β3, KGN and ASU in fibrin scaffold. Immunostained with anti-type II collagen antibodies (a: TGF-β3, b: KGN c: ASU and d: Control) and anti-type X collagen antibodies (e: TGF-β3, f: KGN g: ASU and h: Control) in vitro culture, magnification ×40
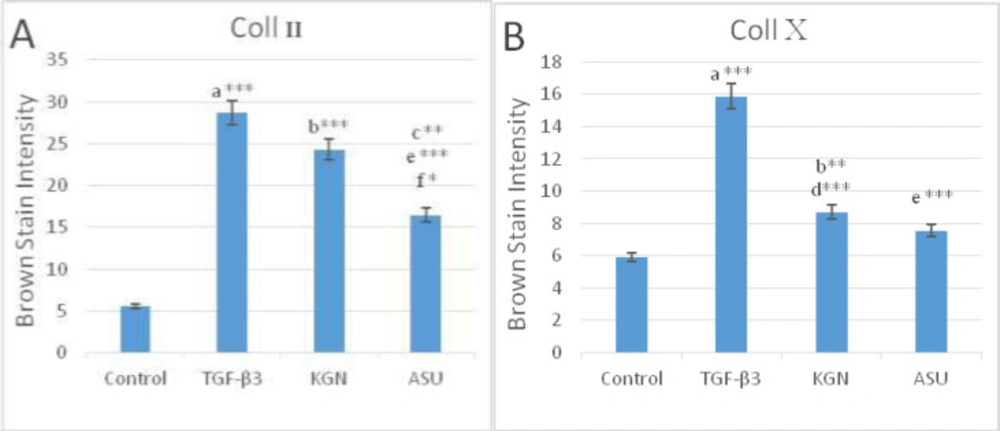
Color intensity demonstrated the deposition of (A) type II and (B) type X collagen protein in control, TGF-β3, KGN and ASU treatment groups. Data are presented as mean ± SD. Error bars represent the standard deviation of the mean. *P < 0.05, **P < 0.01, ***P < 0.001
Histological sections of neo-cartilage formed by TGF-β3, KGN and ASU in fibrin scaffold cultures after 14 days as determined by H&E (a: TGF-β3, b: KGN, c: ASU and d: Control), Toluidine blue (e: TGF-β3, f: KGN, g: ASU and h: Control), Safranin O (i: TGF-β3, j: KGN, k: ASU and m: Control) in the rat model, magnification ×40. Blue arrow: Stem cell with an elongated and fusiform nucleus. Red arrow: Extracellular matrix. Black Arrow: Chondrocytes-like cells inside the lacunae
Immunohistochemical sections of neo-cartilage formed by TGF-β3, KGN and ASU in fibrin scaffold cultures after 14 days. Immunostained with anti-type II collagen antibodies (a: TGF-β3, b: KGN, c: ASU and d: Control) and anti-type X collagen antibodies (e: TGF-β3, f: KGN, g: ASU and h: Control) in the rat model, magnification ×40
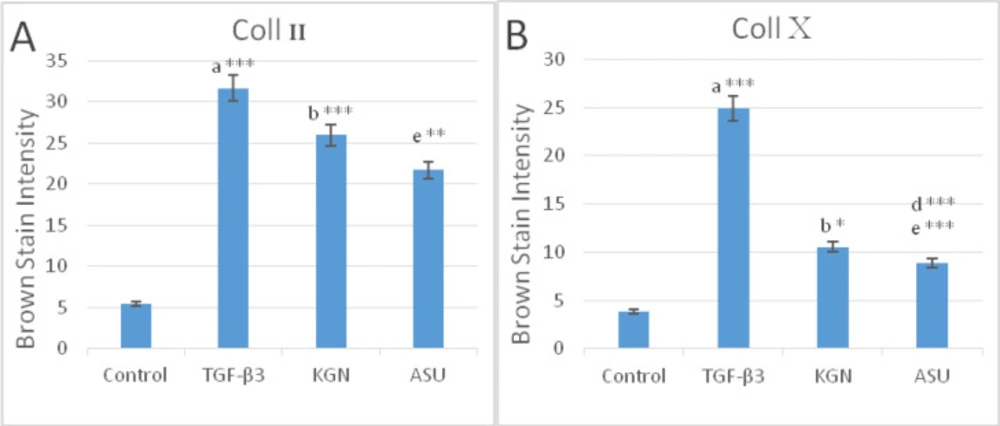
Color intensity demonstrated the deposition of (A) type II and (B) type X collagen protein in control, TGF-β3, KGN, and ASU treatment groups in the rat model. Data are presented as mean ± SD. Error bars represent the standard deviation of the mean. *P < 0.05, **P < 0.01, ***P < 0.001
| Gene | Primer sequences (forward and reverse) |
|---|
| collagen II-F | CTGGTGATGATGGTGAAG |
| collagen II –R | CCTGGATAACCTCTGTGA |
| sox-9 –F | TTCAGCAGCCAATAAGTG |
| sox-9 –R | TTCAGCAGCCAATAAGTG |
| collagen x –F | AGAATCCATCTGAGAATATGC |
| collagen x – R | CCTCTTACTGCTATACCTTTAC |
| Aggrecan-F | GTGGGACTGAAGTTCTTG |
| Aggrecan-R | GTTGTCATGGTCTGAAGTT |
| GAPDH-F | AAGCTCATTTCCTGGTATG |
| GAPDH-R | CTTCCTCTTGTGCTCTTG |