Lapachol Enhances Wound Healing and Sirt1/Sirt3 Expression in Mice
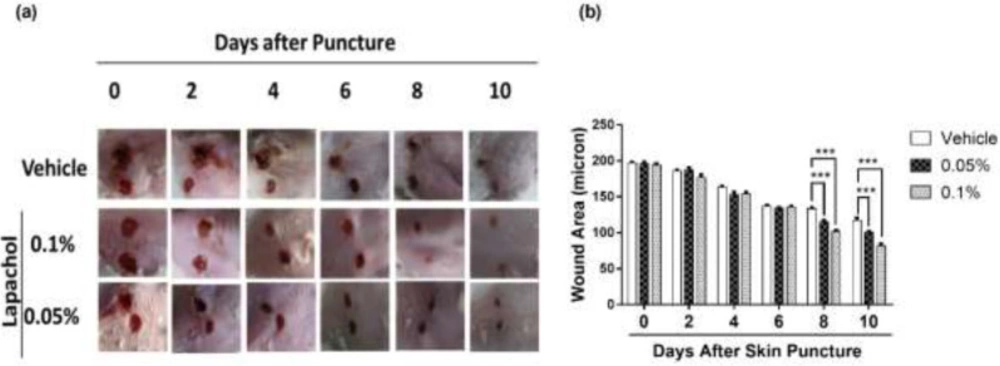
By looking at the photographs from 10 consecutive days, we found that the rate of wound closure was significantly higher in mice treated with lapachol as compared to the mice treated with vehicle only (
Figure 1A). This difference in the rate of wound closure was prominent from day 6 onwards. There was a significant reduction of wound size in lapachol treated mice on days 8 and 10 of treatment compared to the vehicle (
Figure 1A). The difference between the wound size of lapachol treated mice in comparison to a vehicle can be seen clearly in the figure. Moreover, the difference was analyzed statistically and it proved to be significantly higher than the vehicle-treated group (
Figure 1B). Mice in the 0.1% lapachol treated group showed the fastest wound healing with complete wound closure on day 10
th of treatment. We found that mice treated with 0.05% and 0.1% lapachol had perfectly healed wounds compared to the mice group treated with the solvent only. Herein, we got the first clue of lapachol being responsible somehow for the wound healing in diabetic mice, because the vehicle group didn’t show the promising result. We carried on our investigation further by performing western blotting.
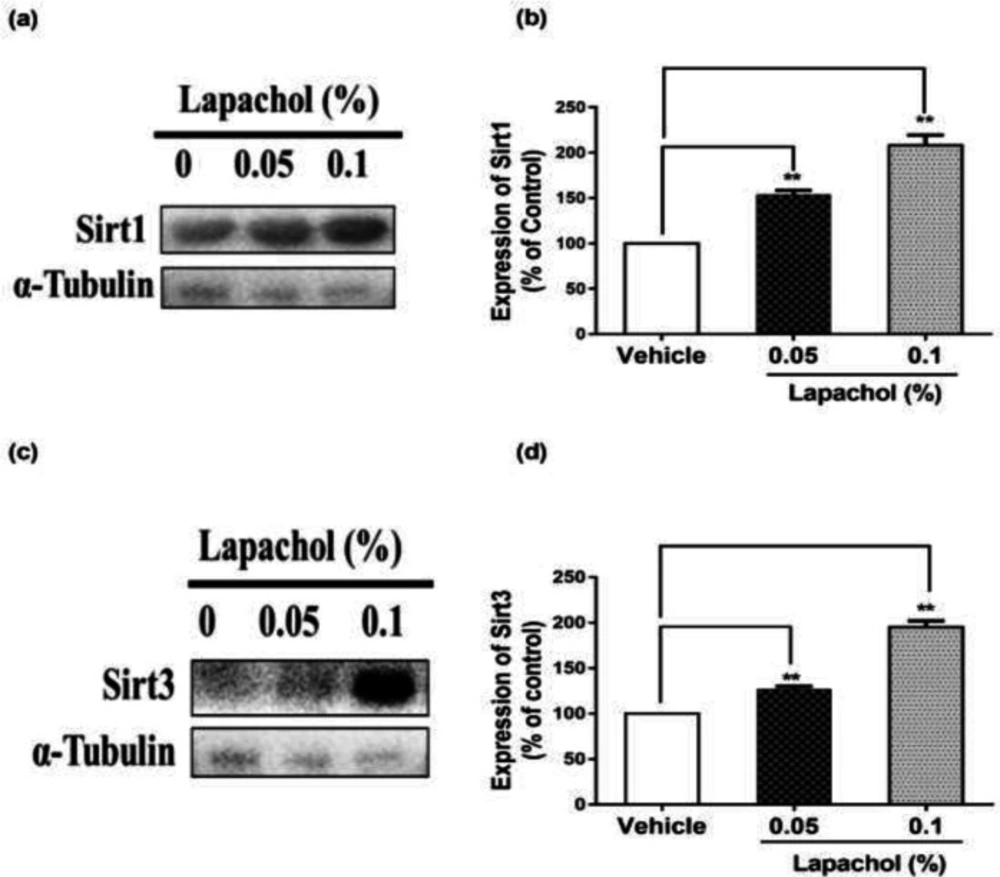
The western blot results showed that sirt1 expression was increased significantly in mice treated with lapachol (
Figure 2A). The diabetic wounded mice treated with solvent only showed low expression of sirt1 as compared to mice treated with lapachol 0.05% and lapachol 0.1%. There was a significant difference between sirt1 expressions in the vehicle as compared to the lapachol treated groups (
Figure 2B). Mice treated with a 0.1% dose of lapachol showed the highest sirt1 expression followed by mice treated with 0.05% lapachol. Similarly, the sirt3 expression in lapachol treated groups was high than the vehicle-treated group (
Figure 2C). Mice treated with lapachol 0.1% were found to have more sirt3 expression as compared to mice treated with lapachol 0.05%. Overall, both lapachol treated mice groups showed significantly higher expression of sirt3 as compared to the non-treated group (
Figure 2D). This significantly increased sirt1 and sirt3 expression in lapachol treated mice groups back our hypothesis and provides an important clue for the role of these proteins in diabetic wound healing.
Lapachol Increases the Expression of Migration and Angiogenesis-Related Proteins in Diabetic Mice Skin
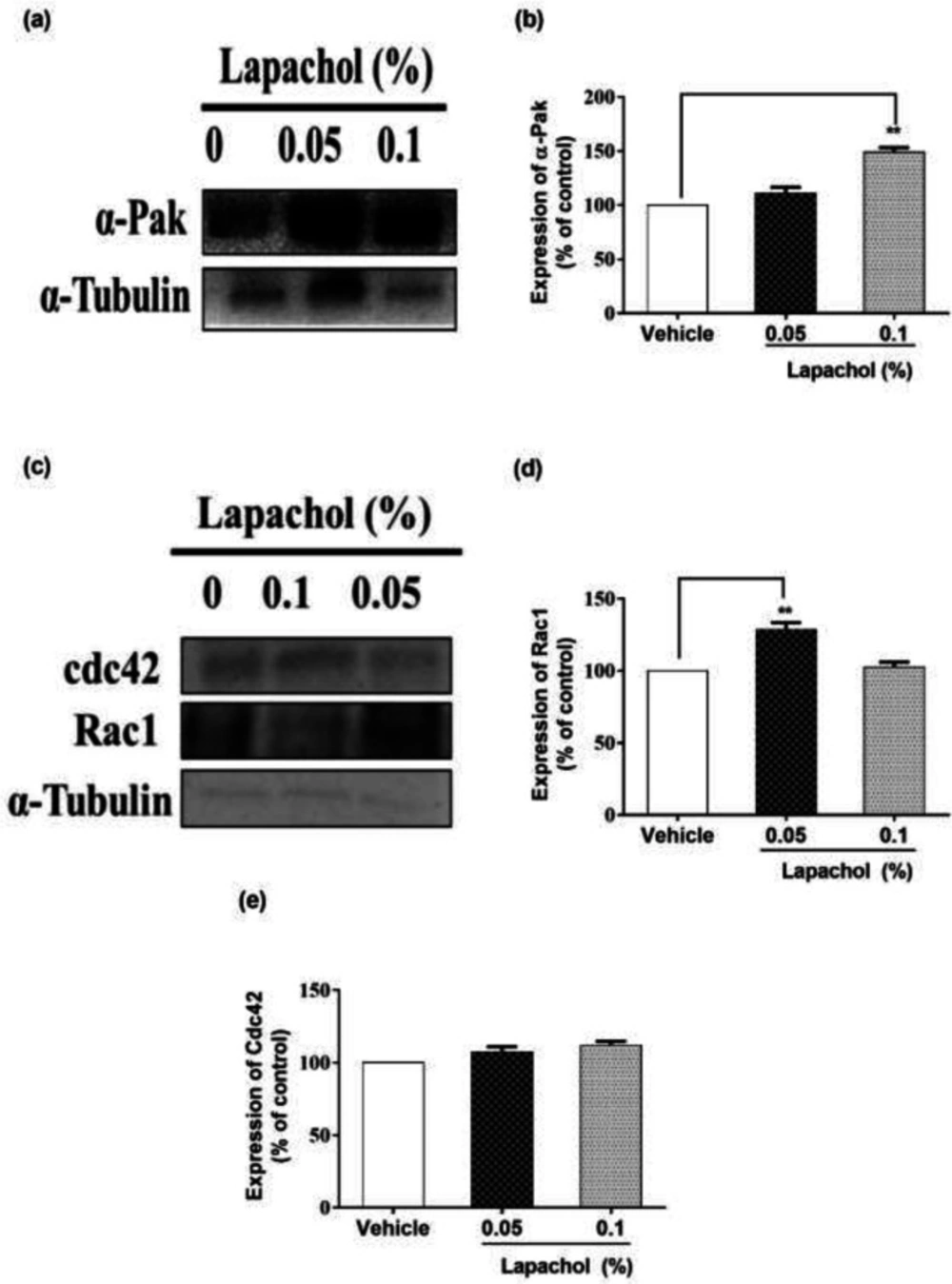
We checked the expression of α-PAK protein by western blotting and it turned to be higher in mice treated with lapachol as compared to the mice treated with vehicle only. Mice treated with lapachol 0.1% showed significantly high expression of α-PAK (
Figure 3A). However, the expression of α-PAK in mice treated with lapachol 0.05% was although higher as compared to the vehicle group, but the difference did not prove to be significant when analyzed statistically. (
Figure 3B).
Similarly, we found that the expression of RAC1 protein was high in lapachol treated mice skin samples as compared to vehicle-treated mice skin samples. There was no significant difference between the expressions of RAC1 in mice treated with lapachol 0.1% and vehicle group (
Figure 3C). However, the expression of RAC1 was significantly increased in mice treated with lapachol 0.05% as compared to vehicle (
Figure 3D), confirming the role of lapachol in wound healing. Next, we ought to check the expression pattern of CDC42 in mice skin samples by western blotting. This time, we did not get any satisfactory results. The expression of this protein was although relatively high in lapachol treated group, specifically the lapachol 0.1% group, but the statistical analysis did not show any significant difference in comparison to the vehicle (
Figure 3E). This part of the experiment needs further investigation and improvement.
Lapachol Enhances Skin Regeneration by Increased Growth Factor Release in Diabetic Mice
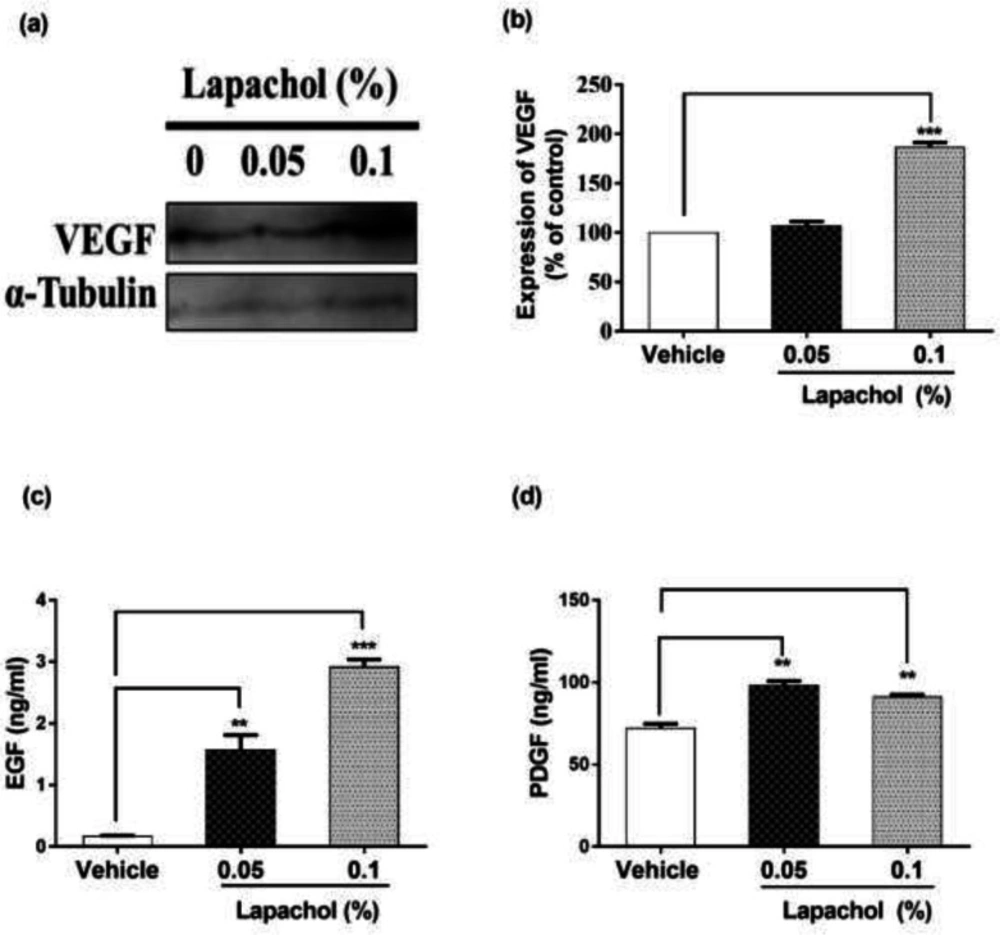
The expression of VEGF was high in lapachol treated mice skin samples as compared to vehicle-treated mice skin samples (
Figure 4A). There was a significant difference between expressions of VEGF protein in mice treated with lapachol 0.1% as compared to the mice treated with vehicle. However, no significant difference was observed between the expression pattern of VEGF in mice skin treated with 0.05% lapachol and mice treated with vehicle. But overall, lapachol treatment resulted in high expression of VEGF, supporting our claim of lapachol as an enhancer of migratory and angiogenic proteins, thus ameliorates diabetic wound healing (
Figure 4B).
We performed ELISA for detecting the concentration of growth factors EGF and PDGF to further evaluate the wound healing potential of lapachol. Skin sample from five mice per group was collected for the experiment. Total cell protein was extracted from each mice group. There was a marked increase in the concentration of EGF in mice treated with lapachol as compared to the group treated with vehicle only. The concentration of EGF was significantly high in mice skin treated with lapachol 0.05% and lapachol 0.1% as compared to vehicle. Mice treated with lapachol 0.1% showed the highest concentration of EGF compared to vehicles. Mice skin obtained from the group treated with vehicle only gave very less concentration of EGF, almost negligible (
Figure 4C). Similarly, lapachol treated mice showed a high concentration of PDGF as compared to the mice treated with vehicle only. There was a significant increase in the concentration of PDGF in lapachol treated groups (
Figure 4D).
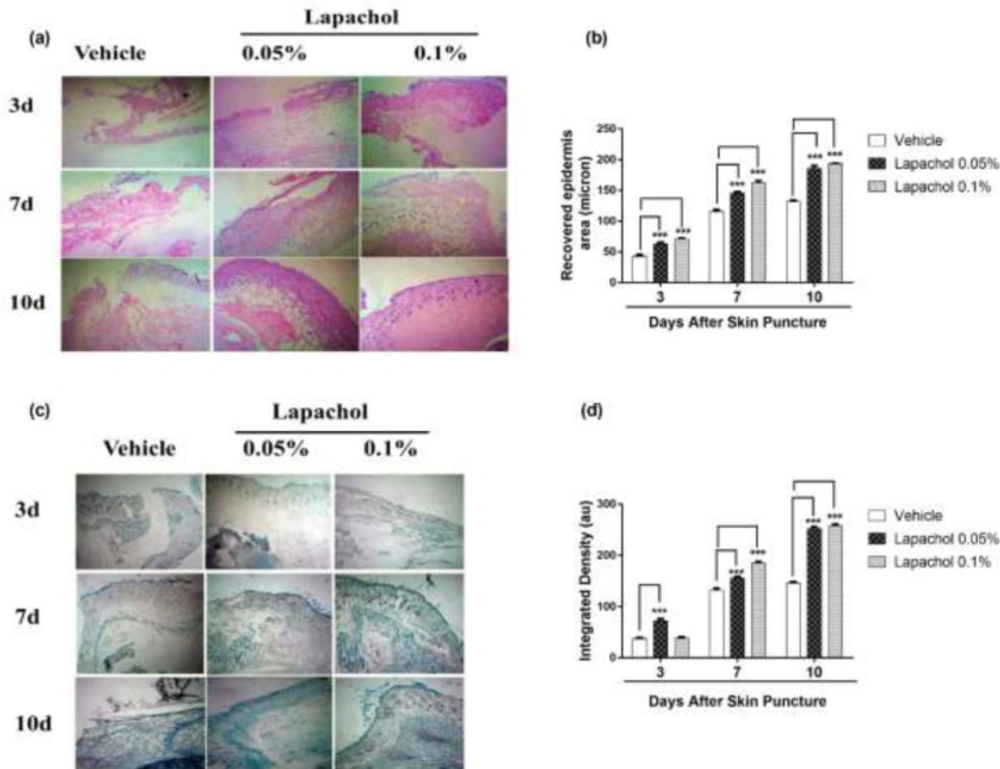
Skin samples collected from 5 mice per group on days 3
rd, 7
th and 10
th of the experiment were stained with Hematoxylin and Eosin (H&E) to check the effect of lapachol on skin regeneration, granulation tissue formation, and neuroepithelium formation. There was faster tissue regeneration and granulation tissue formation in mice treated with lapachol as compared to the vehicle-treated mice on day3, day7 and day 10. Both the dermal and epidermal layers were fully formed and well-characterized (
Figure 5A). The vehicle group treated with solvent only didn’t show remarkable tissue regeneration, also there was no well-characterized formation of epidermal and dermal layers. Lapachol 0.1% treated mice showed a faster wound healing rate with a fully developed epidermal layer on day 7
th of treatment as compared to the 0.05% lapachol. However, both the lapachol 0.1% and 0.05% proved to have the same wound healing effect on day 10
th. There was no significant difference between the two doses on day 10
th, although they were significantly good in comparison to the vehicle on all three-time points (
Figure 5B). The results obtained after H&E analysis strengthened our idea of the potential role of lapachol in diabetic wound healing, as skin regeneration can be observed (
Figure 5A).
Further, we aimed to investigate the effect of different concentrations of lapachol on collagen deposition and organization. For this purpose, mice skin samples from all three groups were collected and stained with Masson’s trichrome stain. Intriguingly, the same results were obtained as stated previously. Mice treated with lapachol showed more collagen deposition shown in blue color with uniform and well-organized collagen as compared to the group of mice treated with vehicle only (
Figure 5C). The amount of collagen was not only less in the vehicle-treated group but it was also haphazardly organized on both day 7 and day 10 post-injury. There was a significant difference between collagen deposition in mice skin treated with lapachol as compared to the vehicle-treated group on days 7
th and 10
th -post-injury. Mice treated with lapachol 0.1% showed more collagen deposition on day 7
th of treatment as compared to mice treated with lapachol 0.05%. However, the effect of both concentrations of lapachol on collagen content was almost the same on day 10
th of treatment (
Figure 5D).