Animals
Albino Wistar rats of both sexes weighing between 250-300 g were acclimatized to laboratory conditions (+20 °C ± 2, 12 h light-dark cycle) for two weeks prior to the experiment, with free access to standard pellet chow and water. With respect to the laboratory analyst and statistical analyst, they were blinded to the groups and treatments allocated to the rats. This study was conducted with the consent of the Animal Ethics Committee of the Marmara University (Approval no: 45.2010 mar).
Surgery and Experimental Design
Four groups of 6 rats were formed in our study. All animals were weighed at the beginning of the experiment. They were then divided into the control group, the BDL group, the SPL1 group and the SPL2 group. All rats were anesthetized (ketamine and 0.75 mg/kg chlorpromazine, i.p.), and the abdomen was opened via a midline laparotomy. In the control group (sham-operated), the rats were incised, the incision was resealed, and only saline (SF) was administered. For the induction of cholestasis (BDL group), the bile duct was doubly ligated at the hepatic hilum. This group was divided into three subgroups; the rats in the SPL1 group received spironolactone (Aldactone tablets, Ali Raif Pharmaceuticals, Istanbul, Turkey) (200 µmol/kg oral, dissolved in 60 mM propylene glycol (
10)) daily for 3 days, starting on the day of the operation. The rats in the SPL2 group received SPL (200 µmol/kg oral, dissolved in 60 mM propylene glycol) daily for three days, starting 3 days after the operation. The BDL group received 60 mM propylene glycol oral (the vehicle) for 3 days. At the end of the experiment, 18 h after the last dose, the animals were decapitated and blood and tissue (liver) samples were collected. Tissue samples were stored in formalin for histological examination and at -80 °C for biochemical evaluations.
Liver function tests
As the markers of hepatic function and tissue injury, the activities of alanine aminotransferase (ALT), aspartate aminotransferase (AST) and alkaline phosphatase (ALP) enzymes, and the total and direct bilirubin (TBIL, DBIL) concentrations were studied with an automated analyzer (Bayer OpeRA, Bayer Corp., Germany) using routine analysis kits (respectively REF 80027, REF 92025, REF80014, REF 80403, Biolabo Europe S.A., Maizy, France).
Measurement of Serum Cytokines Concentrations
Serum NF-ĸB, TNF-α and IL-6 concentrations quantified according to manufacturer's instructions and guidelines using rat-specific commercially available ELISA test kits (BioSource Europe S.A.; Nivelles, Belgium).
Determination of tissue Myeloperoxidase (MPO) activity
Determination of the myeloperoxidase (MPO) level in the tissue was made by the Hillegeas method (
11). Immediately after decapitation, the extracted tissues were washed with saline to remove blood and debris, then dried with filter paper and weighed. The tissue samples were homogenized in 10 vol of ice-cold potassium phosphate buffer (50 mM/L K2HPO4 (pH 6) containing hexadecyltrimethylammonium bromide (HETAB; 0.5%, w/v). The homogenate was centrifuged at 30,000g for 10 min at 4 °C, and the supernatant was discarded. The pellet was then rehomogenized with an equivalent volume of 50 mmol//L K2HPO4 containing 0.5% (w/v) HETAB (Sigma-Aldrich, St. Louis, USA) and 10 mmol/L ethylenediaminetetraaceticacid (EDTA, Sigma). MPO activity was assessed by measuring the H2O2-dependent oxidation of o-dianisidine dihydrochloride (Sigma-Aldrich). One unit of enzyme activity was defined as the amount of the MPO present per gram of tissue weight that causes a change in absorbance of 1.0/min at 460 nm and 37 °C.
Chemiluminescence Experiment
The chemiluminescence (CL) of luminol and lucigenin was detected to measure reactive oxygen species in tissues. Measurements were made at room temperature using a Junior LB 9509 luminometer (EG&G Berthold, Germany). The samples were placed into vials containing PBS-HEPES buffer (20 mM HEPES, pH 7.2, 0.5 M PBS). Reactive oxygen species were measured after adding enhancers lucigenin or luminol at 0.2 mM final concentration. Luminol detects reactive species; namely, it is selective for -OH, H2O2, HOCl radicals, and lucigenin is selective for O−2 (
12,
13). Measurements were made at one-minute intervals and results were given as the area under the curve for a measurement duration of 5 min. The results were corrected for wet tissue weight (relative light units (rlu)/mg tissue) (
14).
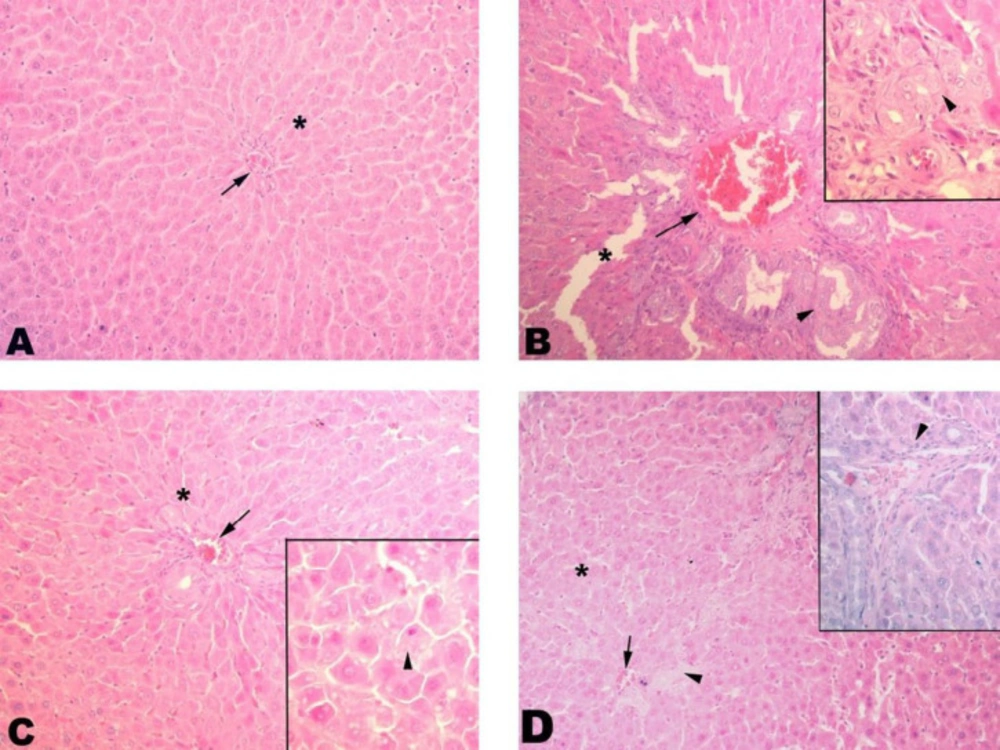
Histopathological Examination
The tissues were washed in tap water for at least 3 h or 1 night after being taken into 10% formalin, and dehydration was performed with increasing alcohol concentrations (15 min with 70% alcohol, 15 min with 90% alcohol, 30 min with 96% alcohol, twice for 30 min with 100% alcohol, and twice for 30 min with 100% toluene). Subsequently, they were kept in paraffin blocks at 60 °C for 1 night and the next day, the tissue was embedded in paraffin blocks. After blocking, sections of 5-6 mm thickness were obtained from the tissues and placed on the slide and left in toluene for 2 h for separating from paraffin, then reduced to water with reduced concentrations of alcohol (2 min with 100% alcohol, 2 min with 90% alcohol, 2 min with 70% alcohol) and left in distilled water, treated with hematoxylin for 15 min, then left for purpling for 10 min in tap water. After applying distilled water with eosin for another 5 min, dehydration was carried out with increasing alcohol concentrations (2 min with 70% alcohol, 2 min with 90% alcohol, 2 min with 96% alcohol, 10 min with 100% alcohol). After this it was washed 2 times with toluene (1st bath for 5 min, 2nd bath for 10 min) and covered with the Entellan before examining under the light microscope.
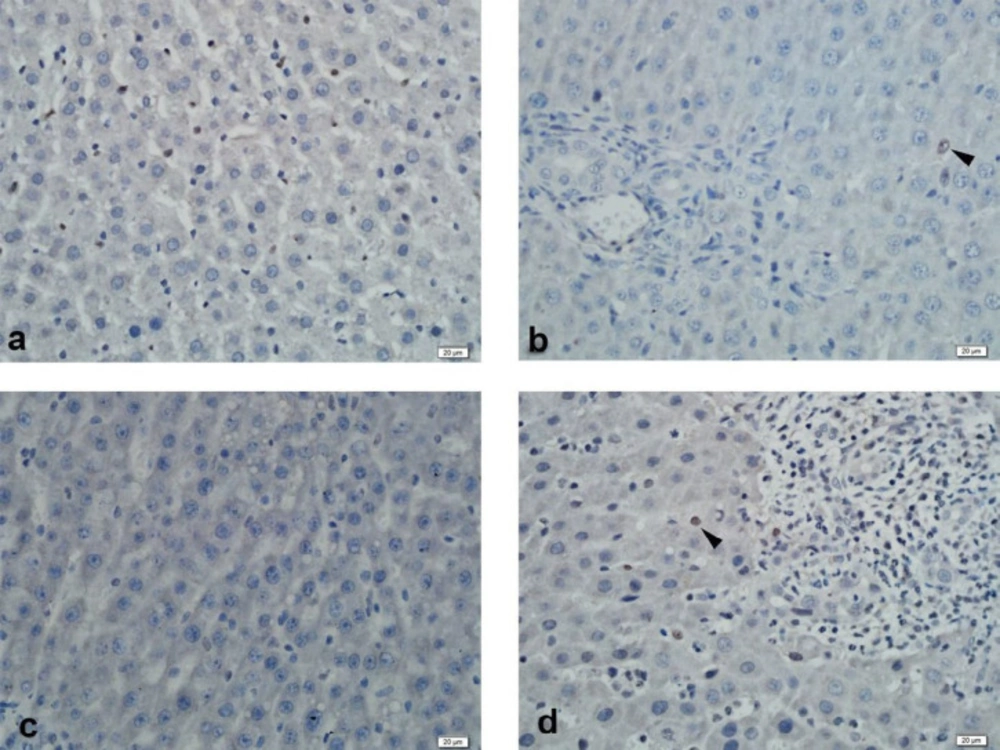
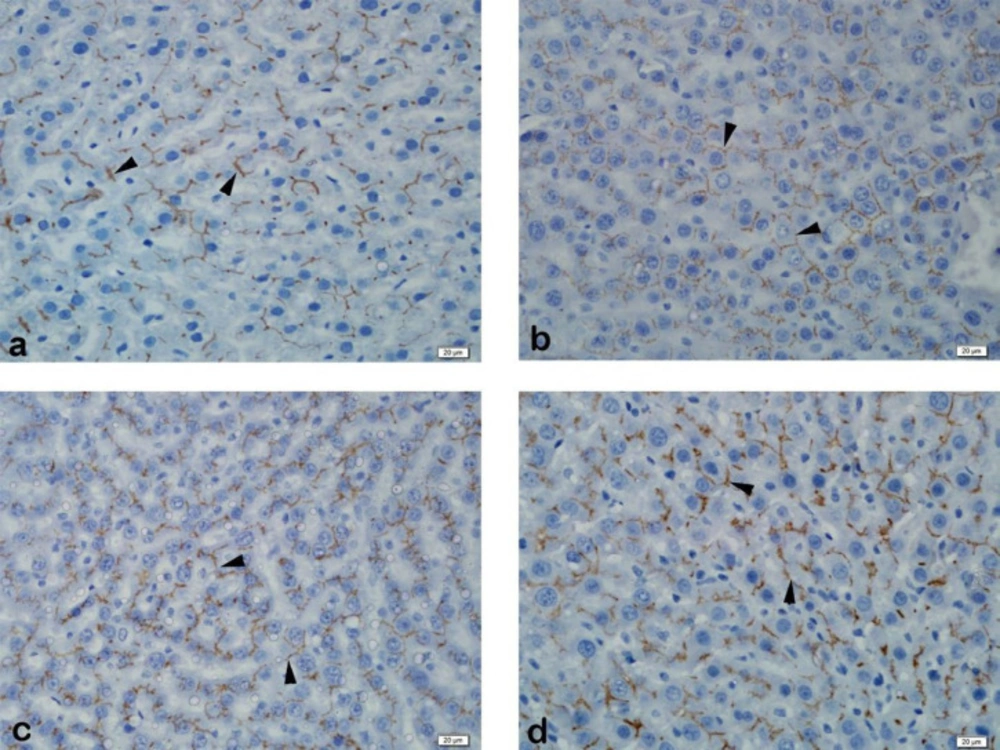
Immunohistochemistry
The Streptavidin-Biotin peroxidase immunohistochemistry staining method was performed to demonstrate immunoexpression of MRP2 and PXR in formalin-fixed paraffin-embedded tissues. In this method, 3μm thick sections were obtained from the paraffin-embedded tissues to positively charged slides and deparaffinized at 37 ºC for 1 night, followed by deparaffinization by keeping in three separate xylenes for 5 min. The sections were passed through two separate 96% ethanol for 10 min, and the endogenous peroxidase activity in the tissue was suppressed with 3% hydrogen peroxide (in methanol) for 20 min. The sections that were washed with distilled water were subjected to an antigen recovery process with a 200W pH 6 citrate buffer solution in a microwave oven to expose the masked antigens. The slides cooled to room temperature was washed with two separate phosphate buffer solutions (PBS) for 5 min and 10 min of protein blocking (Histostain Bulk Kit, Invitrogen LAB-SA Detection System, UK) was carried out to prevent non-specific staining. Following blocking, MRP2 (M2 III-6, Abcam, UK) dilution of 1:100, and PXR (bs-2334R, Bioss, USA) at a dilution of 1:200 was instilled and the specimens were incubated at room temperature for 1 h. This was followed by washing the sections with two separate PBS for 5 min each, and they were placed in biotinylated secondary antibody (Histostain Bulk Kit, Invitrogen LAB-SA Detection System, UK) for 10 min. After washing again with PBS, streptavidin peroxidase (Histostain Bulk Kit, Invitrogen LAB-SA Detection System, UK) was instilled, and they were incubated for 10 min. 3,3’-diaminobenzidine (DAB) chromogen was instilled into the sections that were washed with two separate PBS for five min each, and staining was checked after 5 min of incubation. The sections washed with distilled water were counterstained with Mayer's hematoxylin and dehydrated by passing through ethanol. Tissues that were transferred into xylene and covered with appropriate closure material were evaluated semi-quantitatively with a light microscope (BX51 Olympos, Japan).
Chemicals and reagents
Chemicals and reagents used for the study included SPL (Aldactone tablets, Ali Raif Pharmaceuticals, Istanbul, Turkey), ELISA kits specific for rat cytokine NF-κB (Biosource International, Nivelles, Belgium), HETAB (Sigma-Aldrich, St. Louis, USA), o-dianisidine 2HCl (Sigma-Aldrich), lucigenin (bis-N-methylacridiniumnitrate), and luminol (5-amino-2,3-dihydro-1,4-phthalazinedione (Sigma-Aldrich). Chemicals and reagents used for immunohistochemistry were a serum-blocking solution (Histostain Bulk Kit, Invitrogen LAB-SA Detection System, Paisley, UK) and antibodies to PXR (bs-2334R, Bioss, Salem MA, USA) Mrp2 (M2 III-6, Abcam, UK). All other chemicals used in this study were analytical grade.
Statistical analysis
Statistical analyses were performed using the GraphPad Prism 6.0 (GraphPad Software, San Diego, CA, USA). All findings were expressed as mean ± standard deviation. Statistical analysis was performed with the one-way analysis of variance (ANOVA) and Tukey's test for further analysis. P values lower than 0.05 were considered significant.