
Introduction
Experimental
Results
Discussion
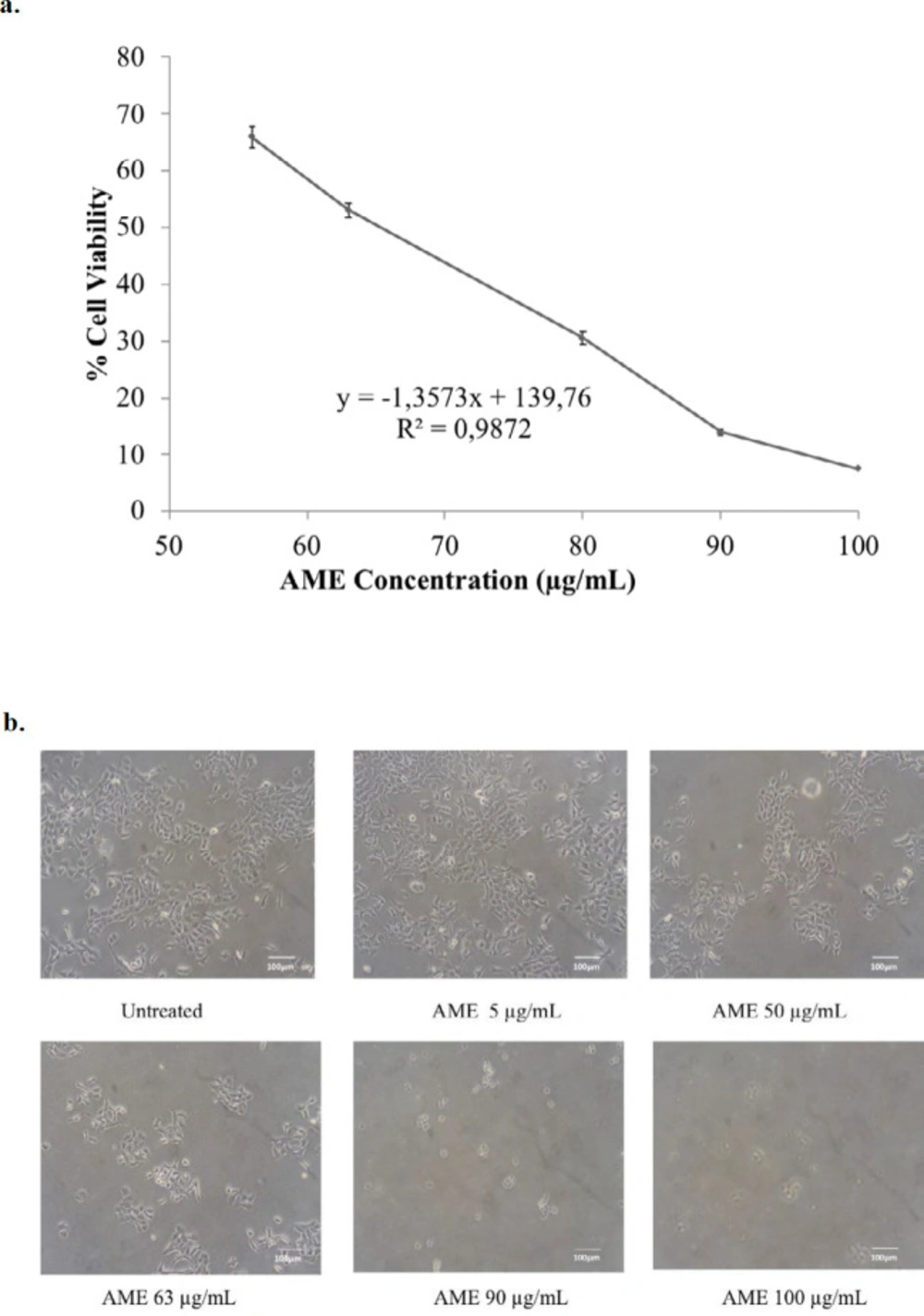
Cytotoxic effect of AME on 4T1 cells. 4T1 cells (5 × 104 cells/mL) were treated with AME for 24 h and then subjected to MTT assay. The viable cells were then quantified using a 450-nm ELISA reader. (a) Percentage of 4T1 cells viability. The viable cells were calculated in accordance with the analysis procedure (p < 0.05). (b) Cells’ appearance after being treated with AME under an inverted microscope with a 100x magnification
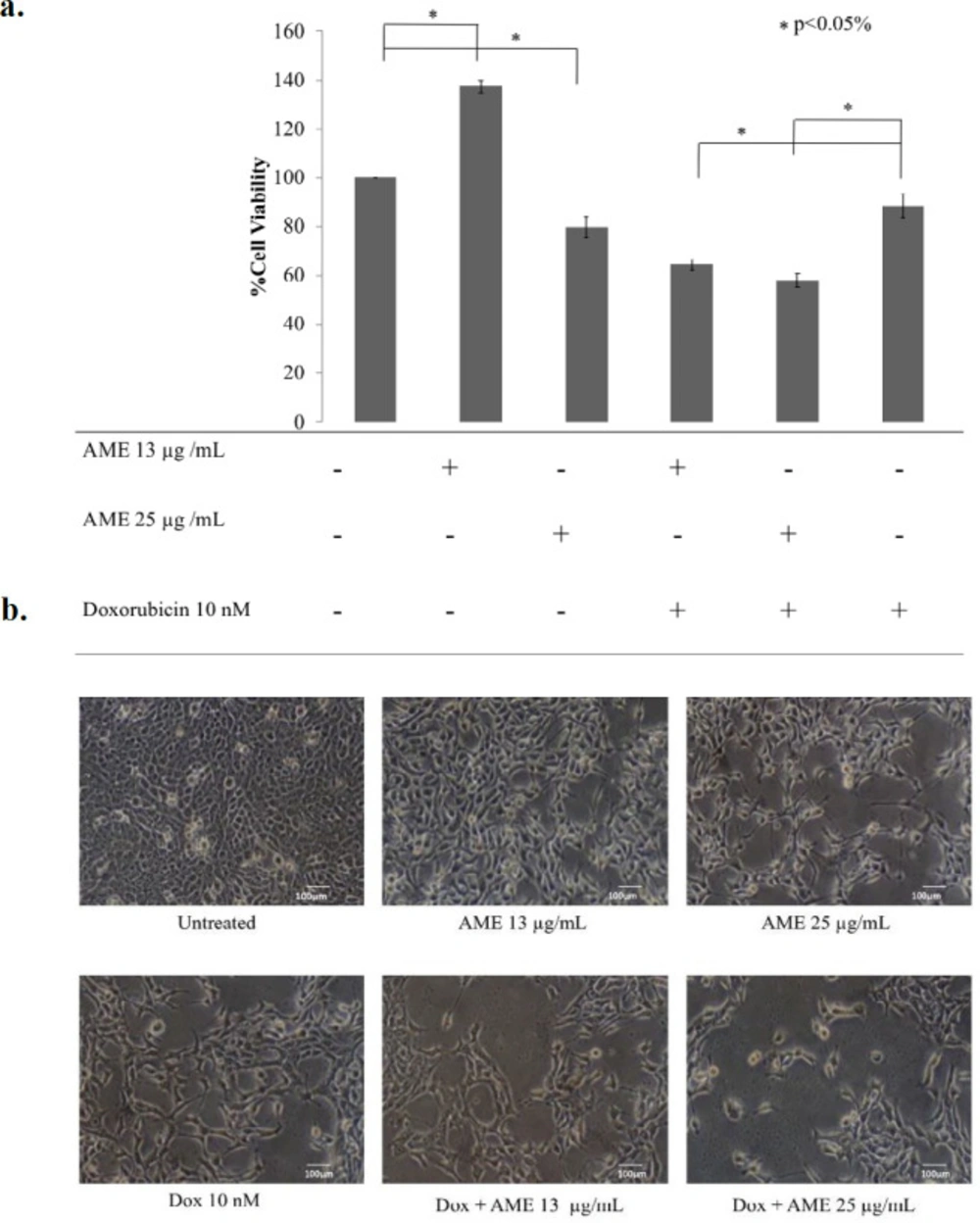
Cytotoxicity of AME in combination with dox. 4T1 cells (5 × 104 cells/mL) were treated with 13 μg/mL and 25 μg/mL of AME, and its combination with dox (10 nM) for 24 hwas then subjected to counting with trypan blue. (a) The percentage of 4T1 cells viability.The viable cells were calculated in accordance with the analysis procedure (p < 0.05). (b) The cells’ appearance after being treated with AME under an inverted microscope with a 100× magnification
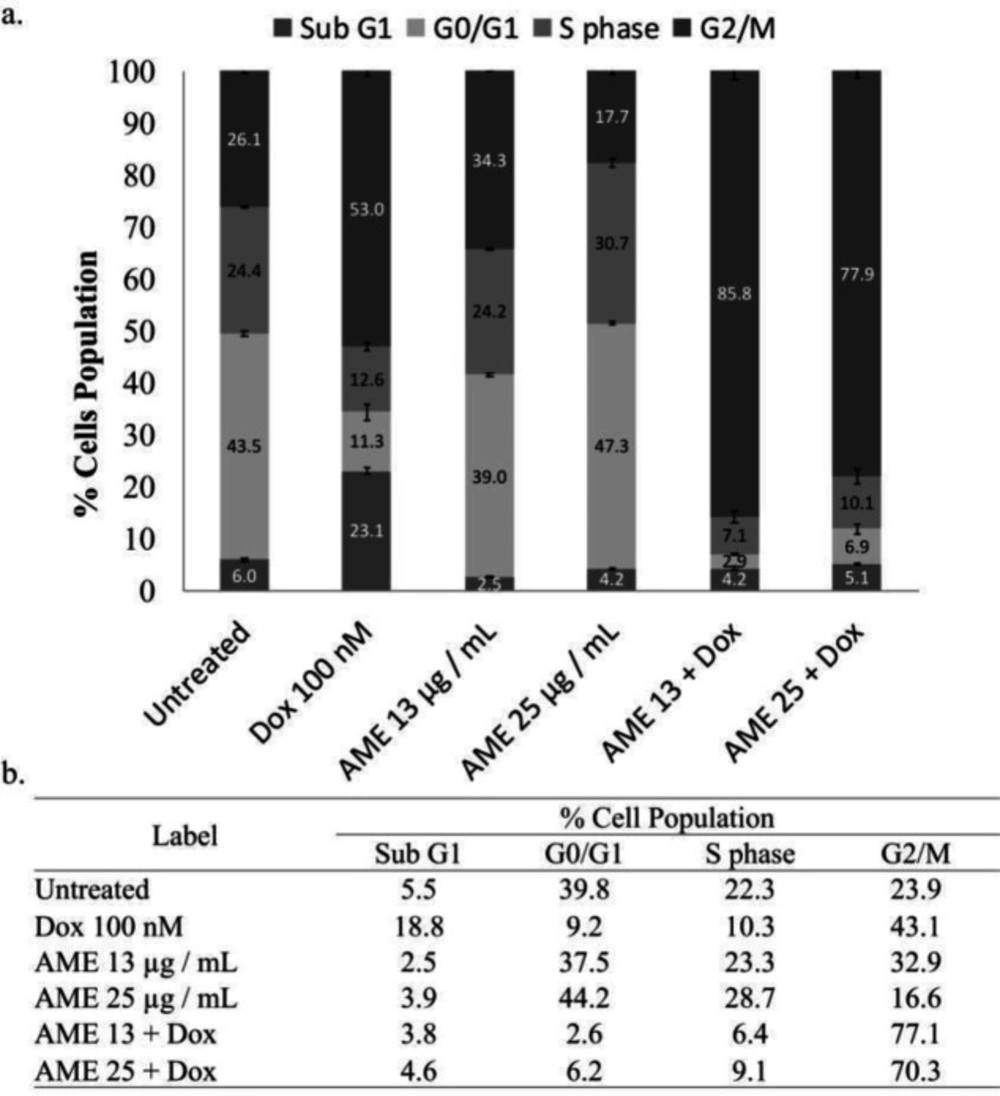
Cell cycle distribution effect of AME alone and in combination with dox. (a) 4T1 cells (3 × 103 cells/mL) were treated with 12.5 and 25 µg/mL of AME alone and in combination with 100-nM dox for 24 h and subjected to cell cycle analysis under PI staining with flow cytometry (n = 3). (b) The summarization of % cell population of 4T1 cells for each phase
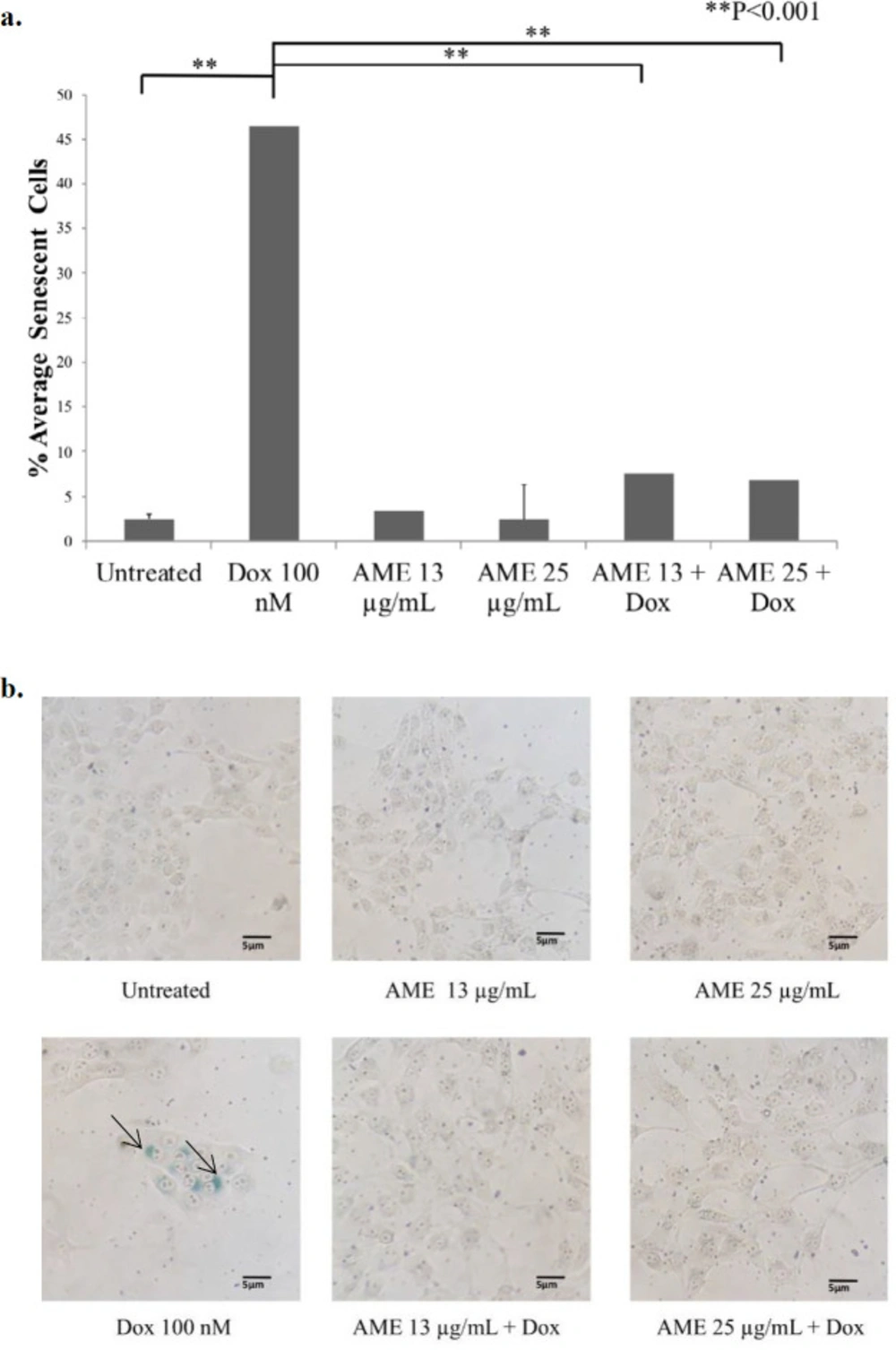
Effects of AME on 4T1 cells in-vitro. (a) 4T1 cells (5 × 105 cells/mL) were treated with Annona muricata (12.5 μg/mL and 25 μg/mL) alone and in combination with dox (100 nM) for 1, 2, and 3 days and subjected to β-galactosidase staining. As a positive control, cells were treated with dox (100 nM) for 3 days. The percentages of senescent cells (β-galactosidase-positive cells) were calculated (n = 3). (b) The senescence induced cells’ appearance after being treated with AME and dox under an inverted microscope with a 100× magnification
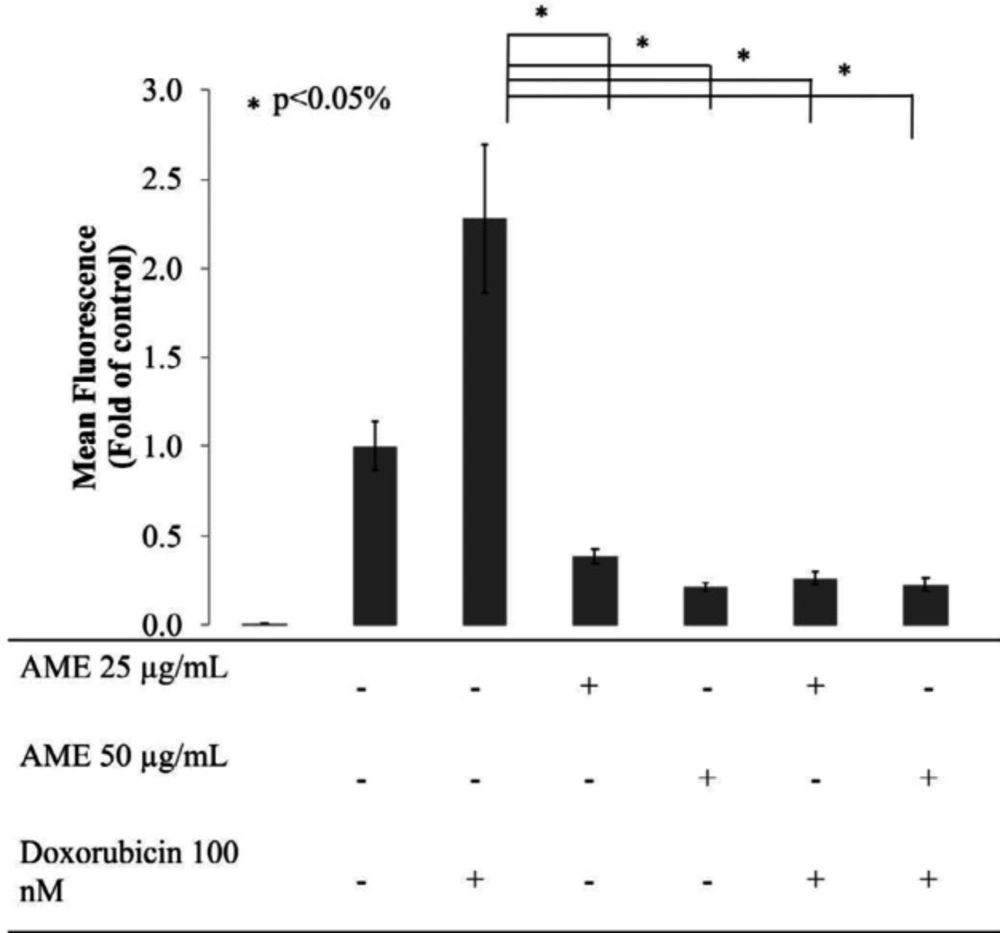
Effect of AME on the intracellular ROS levels of 4T1 cells. Treatment with AME alone and in combination with dox decrease intracellular ROS levels. Cells (8 × 105 cells/mL) were treated with AME (25 and 50 μM), dox (100 nM), and both for 4 h and subjected to ROS detection analysis using flow cytometry. AME decreased the ROS levels in 4T1 cells after the 4-h treatment
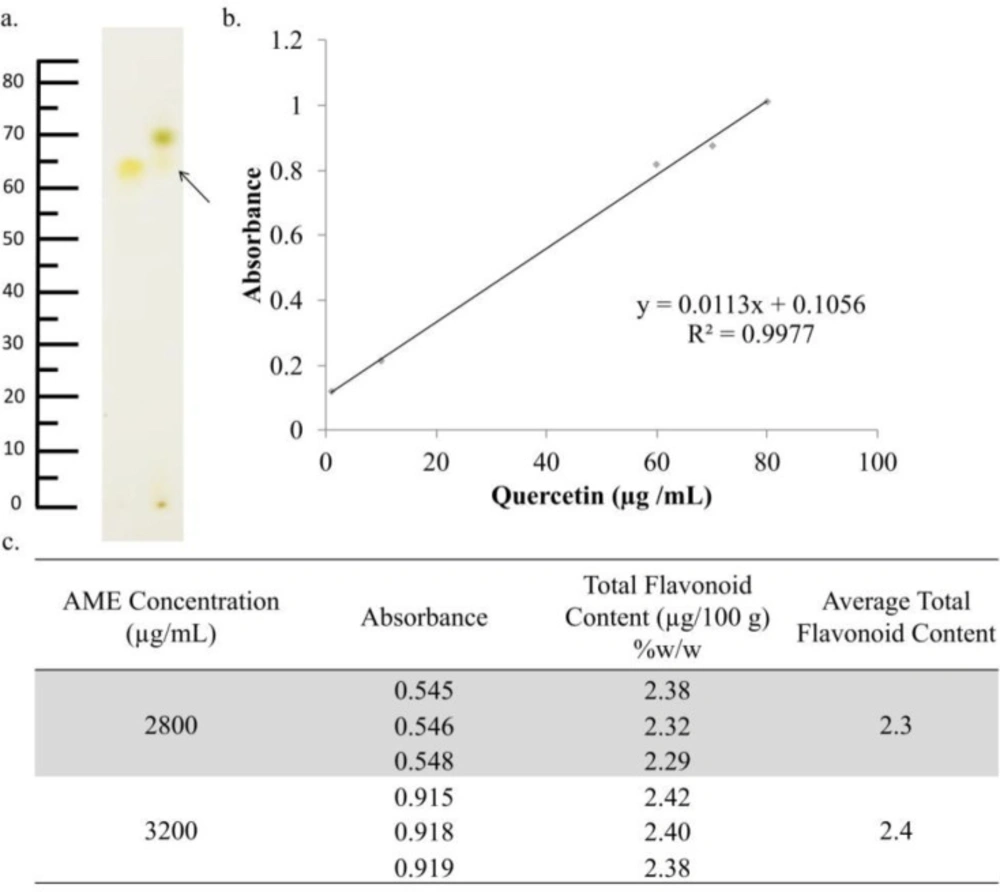
Identification of the chemical content of AME. AME was identified using TLC, while the total flavonoids were determined using spectrophotometry. (a) TLC chromatogram of AME (2) and quercetin (1) showed relatively similar spots between quercetin as standard and AME with an Rf value 0.70. (b) The regression equation obtained from quercetin standard measurement (c) The total flavonoid determination indicated that the total flavonoid content of AME was 2.30% ± 0.05%.
| AME Concentration (µg/mL) | Absorbance | Total Flavonoid Content (µg/100 g) (%) (w/w) | Average Total Flavonoid Content (µg/100 g) (%) (w/w) |
|---|---|---|---|
| 2800 | 0.545 | 2.38 | 2.3 |
| 0.546 | 2.39 | ||
| 0.548 | 2.29 | ||
| 3200 | 0.915 | 2.42 | 2.4 |
| 0.918 | 2.40 | ||
| 0.919 | 2.38 |
| Label | Cell Population (%) | |||
|---|---|---|---|---|
| Sub G1 | G0/G1 | S | G2/M | |
| Untreated | 5.5 | 39.8 | 22.3 | 23.9 |
| Dox 100 nM | 18.8 | 9.2 | 10.3 | 43.1 |
| AME 13 µg / mL | 2.5 | 37.5 | 23.3 | 32.9 |
| AME 25 µg / mL | 3.9 | 44.2 | 28.7 | 16.6 |
| AME 13 + Dox | 3.8 | 2.6 | 6.4 | 77.1 |
| AME 25 + Dox | 4.6 | 6.2 | 9.1 | 70.3 |
