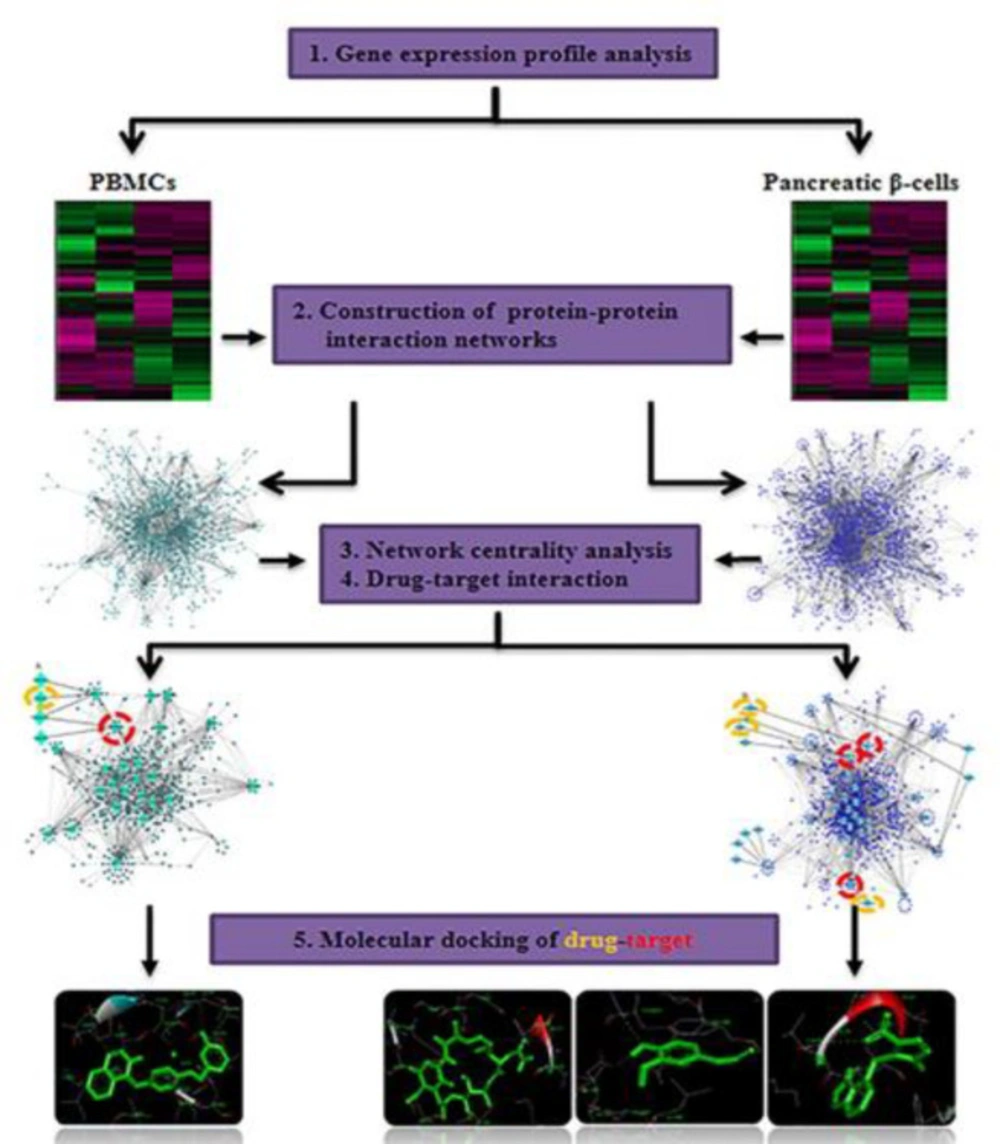
Systems biology approach gives us a comprehensive view to improve our understanding of disease mechanisms and introduce new way for a discovery of novel drugs and repurposing of existing drugs (
35). In this study, differentially expressed genes of PBMCs and pancreatic β-cells in T1D integrated with PPI data. Five centrality parameters, degree, betweenness, closeness, centroid value and eigenvector were measured for nodes in PBMCs and pancreatic β-cells QQPPI networks to find central proteins. A protein is considered a key molecule only if it is selected in at least two centrality measures. By analysis of central proteins and drugs interactions, several candidate drugs and targets were prioritized for T1D. Furthermore, some predicted drug-targets with a relevant to diabetes were used as an input for docking analysis which can calculate the probability of a physical interaction with the given drug and candidate targets.
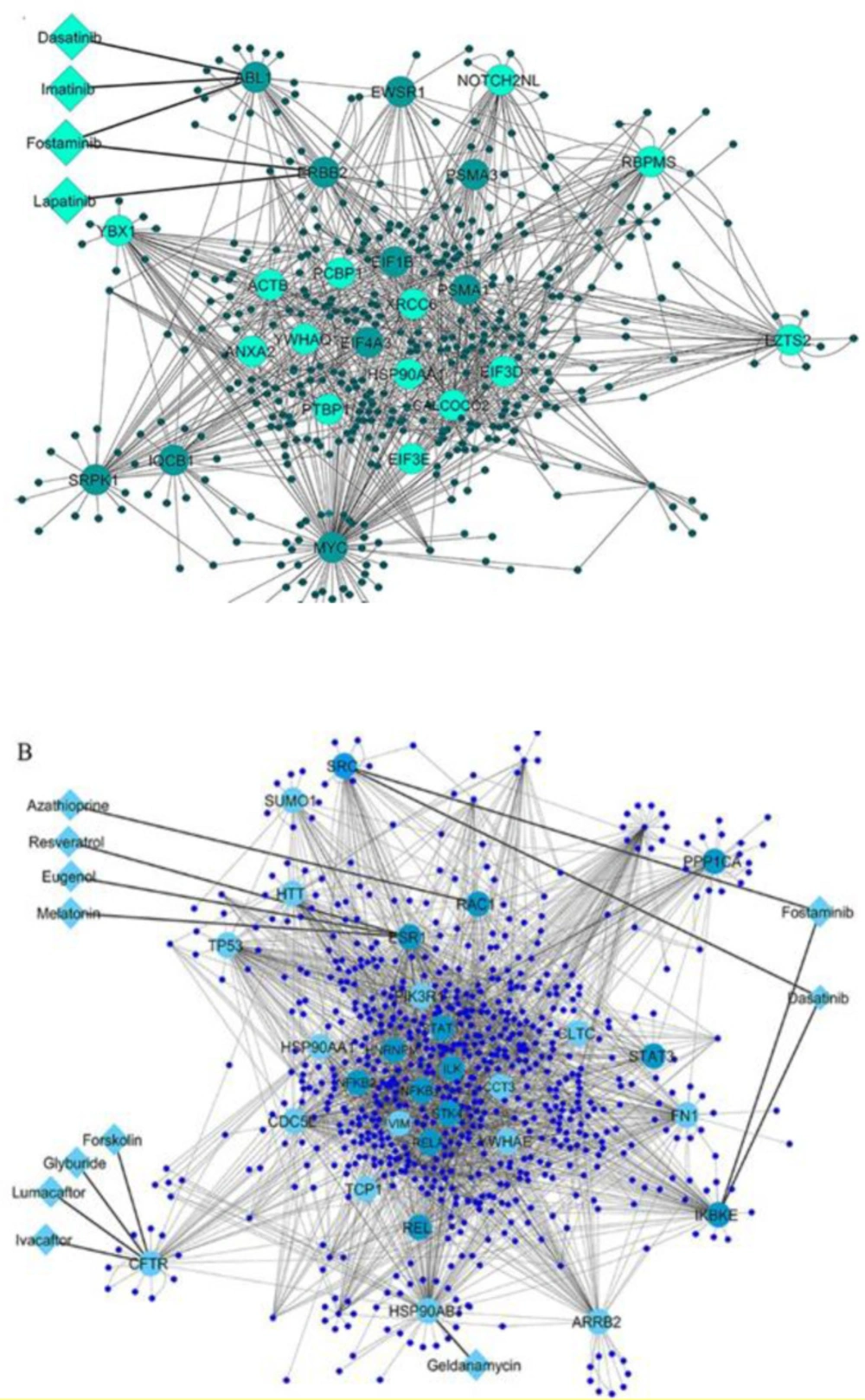
In the current study, fourteen targets and twenty-one candidate drugs were identified. By literature review, thirteen drugs which targeted eight key proteins showed association with diabetes. Some of them such glyburide has been approved for T2D and some ones like imatinib, resveratrol and melatonin are being evaluated in the clinical trials in patients with diabetes. Glyburide targets CFTR gene and it is second-generation sulfonylureas which approved for diabetes management. Glyburide is an antagonist of CFTR. The results of a study revealed a role of CFTR in glucose-induced electrical activities and insulin secretion in β-cells (
36). Grishman
et al. study proposed a glyburide as one of the potential therapies to decrease the progression of T1D due its ability to decrease IL1β levels (
37). Imatinib targets ABL gene. Endoplasmic reticulum stress in β-cell was increased in the NOD mouse as a result of the c-Abl tyrosine kinase activity. Consequently, the unfolded protein response was promoted that ultimately leading to β-cell death; this process might be affected via inhibition of c-Abl by imatinib (
38). Moreover, insulin response was ameliorated in the experimental model of T1D by imatinib. Now, phase II clinical trial of imatinib treatment is ongoing in early-onset patients with T1D (
39). Resveratrol targets ESR1 gene and is being evaluated in the clinical trials in patients with insulin resistance and T2D. Resveratrol binds to estrogen receptors which might be linked to the anti-diabetic effect in diabetes (
40). It is well established that resveratrol diminish blood glucose levels in animals with experimental T1D (
41). In this line, the results of a study demonstrated that resveratrol or 17β-estradiol apparently protected against STZ-induced diabetes in OVX mice; they probably improve antioxidant activities and islet function, promote muscle glucose uptake and prohibit the expression of p-ERK (
42). Melatonin also targets ESR1 gene; it is being assessed in the clinical trials in patients with T2D. Melatonin interferes with estrogen-signaling pathways (
43). The increased level of melatonin synthesis was reported in an animal model of STZ-induced T1D (
44). The improvement of immune response and the anti-inflammatory effect were mentioned as the consequence of melatonin, which might inhibit the disease onset or ameliorate the survival of islet grafts transplanted for T1D therapy (
45).
Moreover, four targets and their candidate drugs that their associations have been proved in the experimental models of diabetes were also prioritized. First, geldanamycin targets HSP90AB1 gene. Geldanamycin is specific HSP90 inhibitor. HSP90 inhibitors such as geldanamycin and its derivatives target HSP90 N terminus and block its ATPase activity; they have been identified as potential treatment strategy in cancer and promising drugs for immune and inflammatory diseases, including diabetes (
46). The result of a study presented that heat shock proteins as well as treatment with geldanamycin noticeably improve diabetic macrophages activation, resulting in compromising mounting of inflammatory and immune responses (
47). Besides, hyperglycemia was reversed by chronic dosing of HSP90 inhibitors in the diabetic db/db mouse model, and insulin sensitivity was made better in the diet-induced obese mouse model of insulin resistance (
32). The second, eugnol targets ESR1. Eugenol is an estrogen receptor antagonist. Several medicinal applications were reported for the eugenol treatment such as antibacterial, antiviral, antioxidant, anti-inflamatory agent (
48). The result of Al-Trad
et al. study showed significant anti-oxidative and anti-inflammatory effect of eugenol in HFD/STZ-induced diabetic rats. Moreover, insulin sensitivity was improved by eugenol and skeletal muscle glucose uptake was stimulated via activation of the GLUT4-AMPK signaling pathway (
33). The third, lapatinib targets ErbB2 gene. Lapatinib is a member of tyrosin kinase inhibitors (TKIs) which can target tyrosine kinase enzymes and interfere with downstream intracellular messaging pathways (
49). Numerous studies represented the glucose-lowering potential of TKIs, which suggest careful attention to apply these drugs to patients with diabetes (
50). Lapatinib is a dual inhibitor of EGFR and ErbB2 receptor tyrosine kinases, by which high glucose-induced apoptosis and vascular dysfunction were refined via resistance to signaling changes influenced by diabetes in the experimental T1D models (
34,
51). The last, azathioprine targets Rac1 gene. Azathioprine suppresses both
T and
B-lymphocyte function (
52). Tiede
et al. reported that azathioprine induces immunosuppression by prohibition of Rac1 activation in T cells, which may clarify the beneficial immunosuppressive effects of azathioprine. Therefore, it might help to design the novel specific therapies for organ transplantation and autoimmune diseases (
53). Furthermore, Veluthakal
et al. identified a known inhibitor of Rac1, NSC23766, which remarkably suppresses reactive oxygen species (ROS) generation in pancreatic islet β-cells in
in-vitro, and significantly inhibits the development of spontaneous diabetes in the NOD mice (
54). Different trials were earlier conducted using azathioprine as an immunosuppressive drug to treat children newly diagnosed with T1D (
52,
55). Although there were partly successful in improving metabolic outcomes in diabetic patients, the reported side effects made it unpopular. However, recently Geliebter
et al. showed the first recent case reports exhibiting the possible positive effect of azathioprine in tertiary prevention of T1D (
56).