Solubilizing of inclusion bodies
To solubilize the inclusion bodies, different concentrations of urea, GdnHCI, and combinations of them were used. As shown in
Figure 1.a, higher amount of hGM-CSF was dissolved when 4 M urea was used as solubilizing agent. Previous studies reported that efficiency of solubilizing with urea or GdnHCI is dependent on the solvent concentration and usually high concentrations of them (
i.e. 6 M GdnHCl and 6-8 M urea) are required to completely solubilize inclusion bodies (
17,
18). However, application of lower concentrations of urea (2-4 M) to solubilize inclusion bodies was also reported by some studies (
19,
20). Similarly, in the present work, solubilizing of inclusion bodies was performed under mild denaturing condition which could preserve secondary structure of the protein, keep the protein in a partially folded state and facilitate the refolding step. Furthermore, high concentrations of the denaturing agent may lead to chemical modifications of protein (
e.g., carbamylation) (
21).
Since solubilizing efficiency of urea is highly pH-dependent (
22), different pH conditions for solubilization of hGM-CSF inclusion bodies were assessed and the optimum pH was found to be 9 (9 ≥ 11 ˃ 5˃ 7) (
Figure 1.b). In the present work, the most efficient solubilizing was observed in 4 M urea at mild alkaline pH, although there are some reports for solubilizing of inclusion bodies using strong alkaline (≥ 12) or acidic (≤ 3) pHs with low concentrations or even no denaturing agents (
23,
24). However, solubilization at extremes of pH may not be an appropriate strategy for the majority of proteins as many of them undergo irreversible modifications such as deamination and acid cleavage (
25).
In addition to pH, the effect of reducing agents (DTT and β-mercaptoethanol) and organic solvents (n-propanol) were evaluated. As revealed by
Figure 1.c, low concentration of β-mercaptoethanol significantly improved the solubilization of hGM-CSF inclusion bodies. This could be explained based on the fact that hGM-CSF molecule has two native disulfide bonds and nine cysteine residues, and this reducing agent can break non-native disulfide bonds and keep cysteine residues of the protein in their reduced state preventing protein misfolding and aggregation (
26). Alternatively, other researchers reported solubilization of hGM-CSF inclusion bodies with 6 M GdnHCl supplemented with 10 mM DTT (
27). However, DTT is more expensive and may interfere with protein purification process (
e.g., immobilized metal affinity chromatography) (
28).
Refolding of inclusion bodies
Among 42 buffer additives, sorbitol, imidazole, Triton X-100, SDS, urea, citric acid, ethanol, and Tricine were selected based on the change in turbidity of samples, densitometry of SDS-PAGE (
Figure 2), and Bradford assay (having the highest protein concentration and the lowest turbidity change).
One of the most important parameters influencing the efficiency of refolding is pH of the refolding solution which determines the net charge of the protein. Proteins usually exhibit an improving solubility below and above of the isoelectric point (pI). Previous studies carried out the refolding of hGM-CSF (pI = 4.95) at a neutral to slightly alkaline pH (7.0-8.0) (
27). Here, 50 mM Tricine, pH = 7 was chosen as the refolding standard buffer based on primary screening of 6 different buffers (Supplementary file,
Table 1). Other seven additives were also evaluated in 30 combination experiments to determine the most important additives that influence the refolding. Three additives including sorbitol, imidazole, and SDS which had the most positive effect were chosen for further optimization
Table 2.
Fifteen experimental runs were designed by Box-Behnken to determine the optimum concentrations of these additives in refolding buffer.
Table 3 shows the analysis of variance for response surface quadratic model describing the correlation between three additives and concentration of refolded hGM-CSF. This correlation could be represented as follows:
Concentration of refolded hGM-CSF (µg/mL) = 117.39 + 4.41 sorbitol + 24.94 SDS + 16.42 imidazole - 3.54 sorbitol×SDS + 25.96 sorbitol×imidazole + 9.33 SDS×imidazole - 36.79 sorbitol2 - 1.56 SDS2 + 12.25 imidazole2
The Model F-value of 65.94 with a very low probability value (< 0.0001)
Table 3 indicates that the model is significant. In adddition, SDS, imidazol, SDS×imidazol, sorbitol
2, SDS
2, and imidazole
2 are signinficant terms (
p < 0.05) in this model. The F-value of 0.64 and probability value of 0.6552 indicates the lack of fit is not significant in the model. As shown in
Table 3, high value of “R-Squared” (0.9916) and “Pred R-Squared” (0.9247) are in resonable agreement with “Adj R-Squared” (0.9766) suggesting a good accuracy of the selected model.
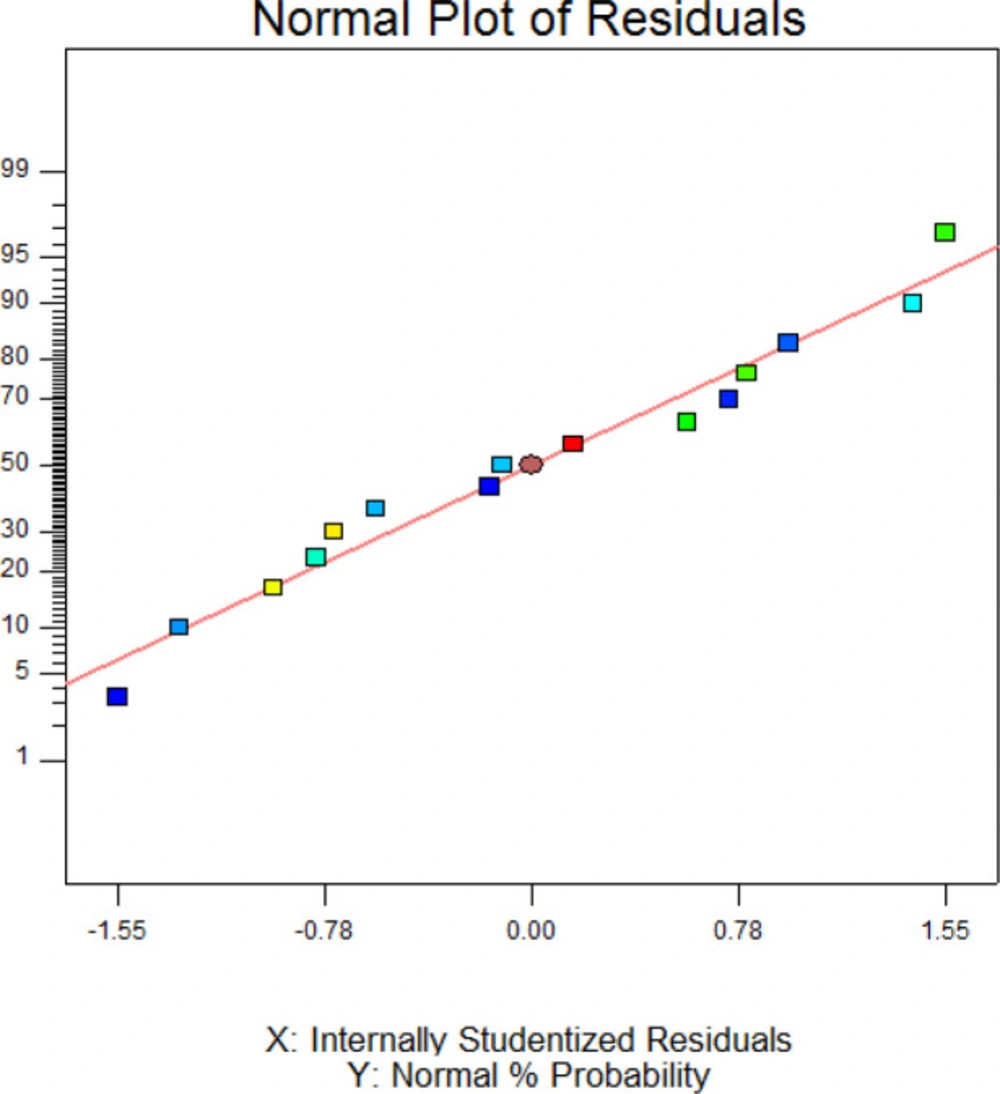
Figure 3 exhibited the normal probability plot of hGM-CSF refolding model. As it is shown, the data points follow a normal distribution which indicates this is a suitable and significant model for predicting hGM-CSF refolding at the different concentarions of these addititves.
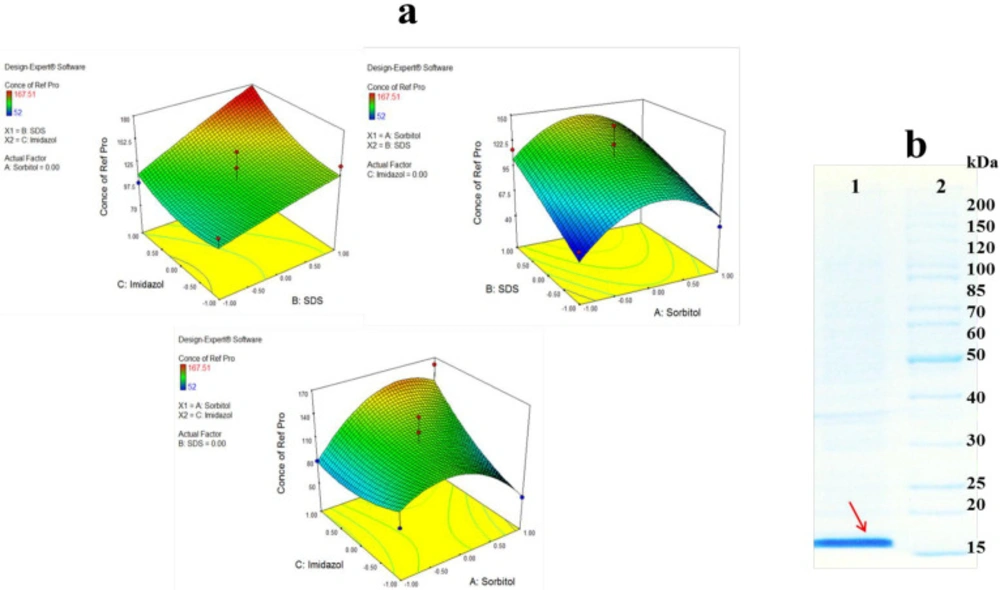
Figure 4.a presents the two factor interactions between three buffer additives. As shown by the 3D surface graphs, refolding yeield of hGM-CSF increased significantly with an increase in SDS concentration, especially in the persence of high concentration of imidazole (
p = 0.0054). Increasing concentrations of SDS at medium value of sorbitol led to an increase in hGM-CSF refolding, althougth the refolding yield reduced as the concentrriaon of sorbitol decreased or increased. SDS is mostly considered as a denaturing agent although in low concentrations (0.1-0.01%) it can decrease misfolding and formation of oligomers after elimination of denaturing agent (
e.g., urea), and during refolding process (
29). In the present study, SDS was used in the presence of sorbitol which is a polyalcohols co-solvent and osmolyte. Michaux
et al., also reported some diol-type solvents can alter denaturing properties of SDS and combination of detergent and co-solvent may help refolding of proteins (
30). Sorbitol can stabilize the protein conformation during refolding process. It was found that the effect of sorbitol on the stability of hGM-CSF was remarkably dependent on its concentration. Similarly, Xie
et al., reported improvement of ribonuclease A stability by sorbitol in a concentration-dependent manner (
31). They proposed general nonspecific bindings of co-solvent to refolded protein are responsible for stabilizing effect of sorbitol. Imidazole as an amino acid derivative can remarkably improve refolding of hGM-CSF. The refolding yield slightly enhanced when the concentration of imidazole increased at low concentration of sorbitol but this effect remarkably increased at high concentration of sorbitol. In consistent with our results, Shi
et al. , reported that imidazole can significantly increase the refolding yield of green florescent protein (GFP). They suggested that imidazole acts like a chaperon molecule and refolding catalyst through enhancement of prolyl isomerization. However, they indicated that the additive is not a protein stabilizer and cannot prevent pH-dependent denaturing of GFP.
Based on DOE results, the optimum buffer for refolding of hGM-CSF was predicted as follow: sorbitol (0.235 M), imidazole (97 mM), and SDS (0.09 %). This optimized condition was used for large scale refolding of hGM-CSF (
Figure 4.b) which yielded a concentration of 180 µg/mL hGM-CSF (corresponds to 18 mg refolded protein from 1 liter of bacterial culture).
Purification
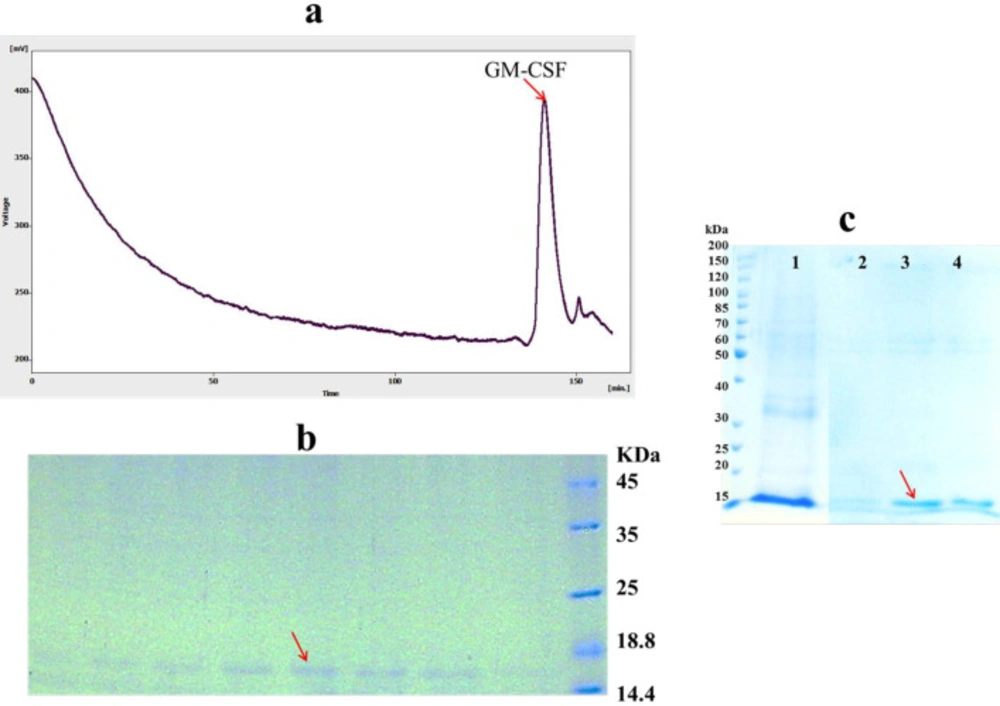
After protein refolding by dilution method, remaining host proteins and aggregated, misfolded and incorrectly folded intermediates of the target protein must be removed to increase the purity of refolded protein (
32). To achieve this, refolded hGM-CSF was further purified by size exclusion chromatography. The refolded hGM-CSF was loaded onto the Superdex 200 column and eluted using a buffer solution of 150 mM NaCl, 50 mM Na
3PO
4, pH = 7.2. A clear peak was observed in the gel filtration profile (
Figure 5.a) eluted in 140-147 min fractions. SDS-PAGE analysis was used to confirm the result of the gel filtration purification of hGM-CSF indicating the purity of eluted hGM-CSF (only one band was observed in 15 kDa region) in the isolated fractions (
Figure 5 b). However, purification of refolded protein by Ni-NTA resin under native condition was not successful (data not shown) possibility due to lower accessibility of hexa histin tag of protein in refolded state (
33).
In addition to direct dilution method, solubilized proteins can be refolded and purified at the same time while bounded to a solid matrix like Ni resin. As shown in
Figure 5.c, high amount of on-column refolded hGM-CSF was eluted in the fraction containing 200 mM imidazole. The purity of the refolded hGM-CSF was 90%. On-column refolding can prevent misfolding and aggregation of protein via decrease in undesirable intermolecular interactions between partially folded monomers (
34).
Analytical and biological characterization of the refolded hGM-CSF
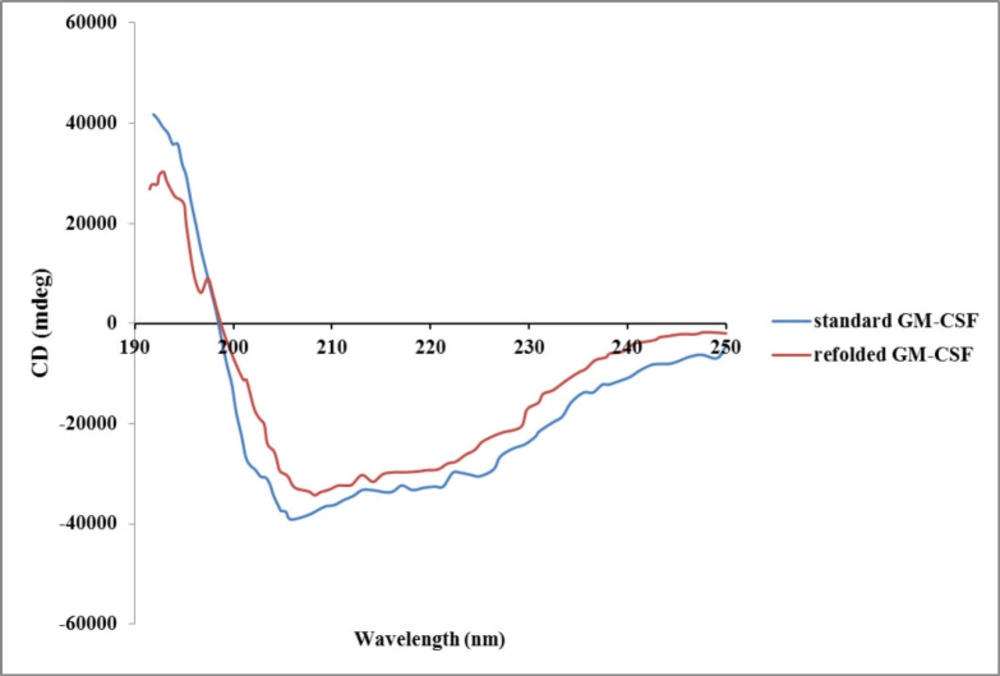
There are different structural- and functional-based methods to monitor refolding of protein. Some structural-based methods like circular dichroism (CD), fluorescence, and nuclear magnetic resonance (NMR) spectroscopies provide valuable information about the folding state (
35,
36). Additionally, functional-based methods such as enzymatic activity measurement and biological assay can be applied to confirm correct recovery of tertiary structure of refolded protein (
37). Crystallography analysis of GM-CSF revealed that it is a highly compact and globular protein consisting of a hydrophobic core. GM-CSF molecule contains four alpha helixes and two beta sheets (
38). In the present study, the secondary structure of the refolded hGM-CSF using optimum buffer condition was evaluated by CD. The CD spectrum of refolded protein was very similar to that of the native protein (
Figure 6) indicating correct formation of secondary structures (
e.g, alpha helix and beta sheet) after refolding.
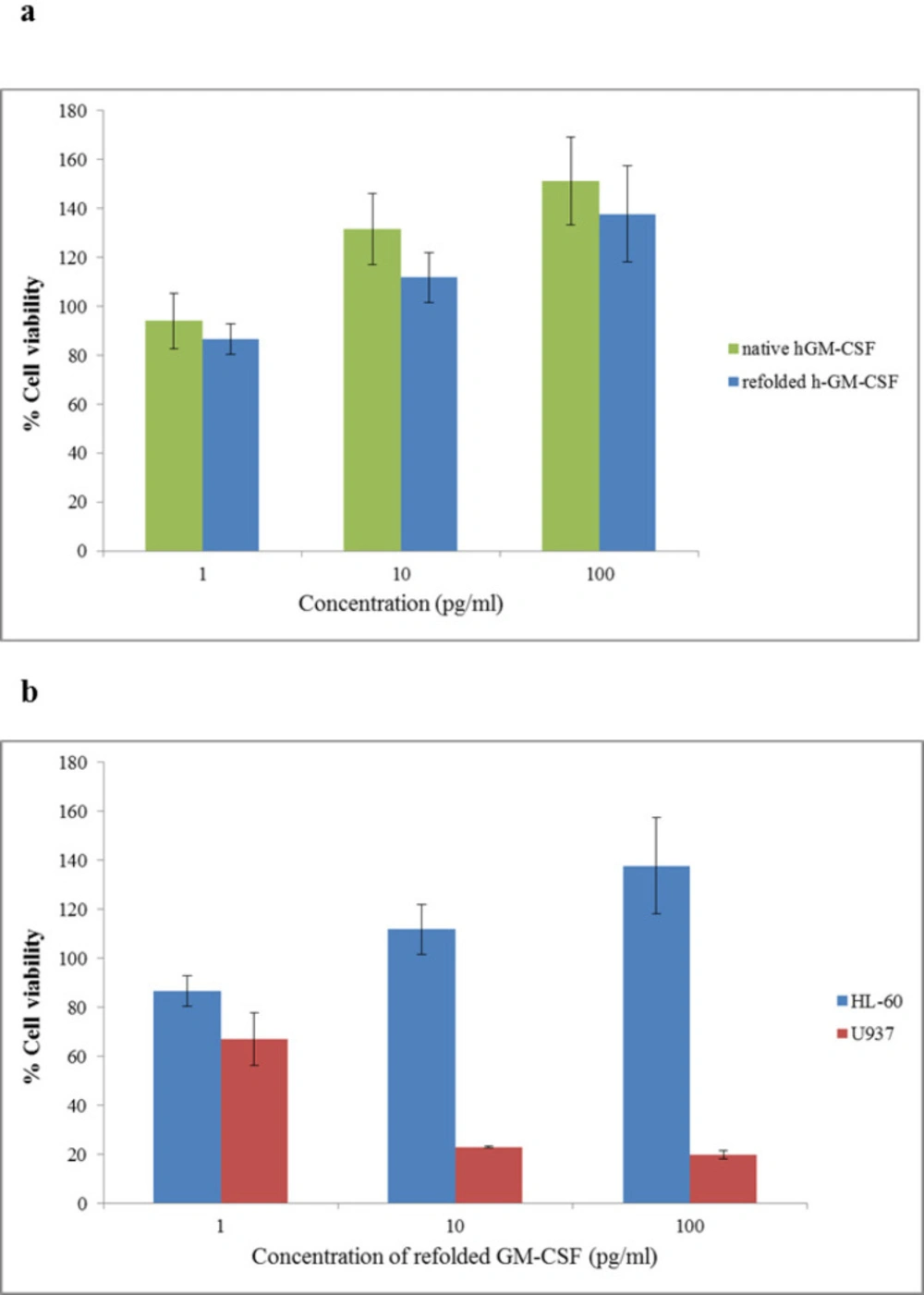
In-vitro biological activity of the refolded hGM-CSF under the optimum condition was evaluated using HL-60 and U937 cell lines.
As it is shown by
Figure 7, refolded hGM-CSF could stimulate proliferation of HL-60 cells in a dose-dependent manner. When HL-60 cells were incubated with 100 pg/mL of the refolded and standard hGM-CSF, cell proliferation increased 37% and 50%, respectively (the difference was not significant;
P ˃ 0.05). Refolded hGM-CSF exhibited similar biological activity to the native protein suggesting successful refolding of the protein. In this study, we evaluated the effect of hGM-CSF on the viability of two hematopoietic cell lines, HL-60 and U937. Interestingly, in contrast to its stimulatory effect on proliferation of HL-60 cells, hGM-CSF significantly inhibited the growth of U937 cells and this inhibitory effect was dose-dependent (
p ˂ 0.05). In agreement with our results, another research group reported inhibitory effect of hGM-CSF on the proliferation of U937 cells (
39). They proposed that secretion of a soluble inhibitory molecule (probably tumor necrosis factor) by U937 cells following treatment with hGM-CSF is responsible for its anti-proliferative effects.
Different yields of hGM-CSF production in
E. coli expression system have been previously reported. Earlier studies achieved a very low yield of protein expression (only 10% of total cell protein) due to toxicity of hGM-CSF to bacterial cell (
40). However, there are some reports on successful application of fusion proteins (
e.g., thioredoxin) for high yield expression of hGM-CSF (
41). Fusion partner removal and further purification steps are needed for this strategy to prevent contamination of the product with a partially cleaved protein. Furthermore, a few additional amino acids are left by most of enzymatic cleavages interfering with bioactivity and immunogenicity of the protein (
42). Therefore, fusion-tag approach is more time-consuming and expensive and it is not a common industrial bioprocessing method particularly at large scales. Alternatively, soluble and native protein can be obtained by refolding inclusion bodies. Thomson
et al. reported successful refolding of hGM-CSF from inclusion bodies and they obtained a yield of 7 mg refolded protein per liter of bacterial culture (
27). In the present study, we applied the systematic optimization of refolding additives which resulted in the highest yield of refolded and bioactive hGM-CSF from inclusion bodies (18 mg from 1 liter of culture) to date. The procedure developed in this work can be applicable to improve the refolding of other proteins with similar structural features.
| Concentration of refolded hGM-CSF µ(g/mL) | Tricine(mM) | Imidazole or C (mM) | SDS or B (%) | Sorbitol or A (M) | |
|---|
| 146.5 | 50 | 50 | 0.05 | 0.25 | Run 1 |
| 160.65 | 50 | 100 | 0.05 | 0.5 | Run 2 |
| 127.42 | 50 | 50 | 0.05 | 0.25 | Run 3 |
| 99.27 | 50 | 100 | 0 | 0.25 | Run 4 |
| 107.33 | 50 | 0 | 0 | 0.25 | Run 5 |
| 167.51 | 50 | 100 | 0.1 | 0.25 | Run 6 |
| 53.66 | 50 | 0 | 0.05 | 0.5 | Run 7 |
| 55.88 | 50 | 50 | 0 | 0 | Run 8 |
| 80.12 | 50 | 100 | 0.05 | 0 | Run 9 |
| 138.24 | 50 | 0 | 0.1 | 0.25 | Run 10 |
| 52 | 50 | 50 | 0 | 0.5 | Run 11 |
| 95.12 | 50 | 50 | 0.1 | 0.5 | Run 12 |
| 113.17 | 50 | 50 | 0.1 | 0 | Run 13 |
| 78.26 | 50 | 50 | 0.05 | 0.25 | Run 14 |
| 76.97 | 50 | 0 | 0.05 | 0 | Run 15 |
| Buffer additive | Stdized effect | Sum of squares | % of contribution | Significance |
|---|
| Sorbitol | 9.2 | 12.03 | 0.23 | 2 |
| Imidazole | 5.40 | 15.81 | 0.30 | 3 |
| Triton | -4.48 | 106.83 | 2.00 | 6 |
| SDS | 4.90 | 492.43 | 9.21 | 5 |
| Urea | 2.79 | 296.14 | 5.54 | 7 |
| Citric acid | -23.29 | 2011.35 | 37.63 | 1 |
| Ethanol | -5.30 | 379.36 | 7.10 | 4 |
| Source | Sum of squares | df | Mean square | F value | Prob > F | |
|---|
| Model | 0.073 | 9 | 8.146E-003 | 65.94 | 0.0001 | Significant |
| A-Sorbitol | 1.531E-004 | 1 | 1.531E-004 | 1.24 | 0.3162 | Not significant |
| B-SDS | 0.057 | 1 | 0.057 | 465.08 | < 0.0001 | Significant |
| C-Imidazole | 7.381E-003 | 1 | 7.381E-003 | 59.74 | 0.0006 | Significant |
| AB | 9.000E-006 | 1 | 9.000E-006 | 0.073 | 0.7980 | Not significant |
| AC | 5.062E-004 | 1 | 5.062E-004 | 4.10 | 0.0988 | Not significant |
| BC | 2.704E-003 | 1 | 2.704E-003 | 21.89 | 0.0054 | Significant |
| A2 | 2.285E-003 | 1 | 2.285E-003 | 18.49 | 0.0077 | Significant |
| B2 | 2.061E-003 | 1 | 2.061E-003 | 16.68 | 0.0095 | Significant |
| C2 | 1.533E-003 | 1 | 1.533E-003 | 12.41 | 0.0169 | Significant |
| Residual | 6.178E-004 | 5 | 1.236E-004 | | | |
| Lack of | 3.038E-004 | 3 | 1.013E-004 | 0.64 | 0.6552 | Not significant |
| FitPure Error | 3.140E-004 | 2 | 1.570E-004 | | | |
| Cor Total | 0.074 | 14 | | | | |
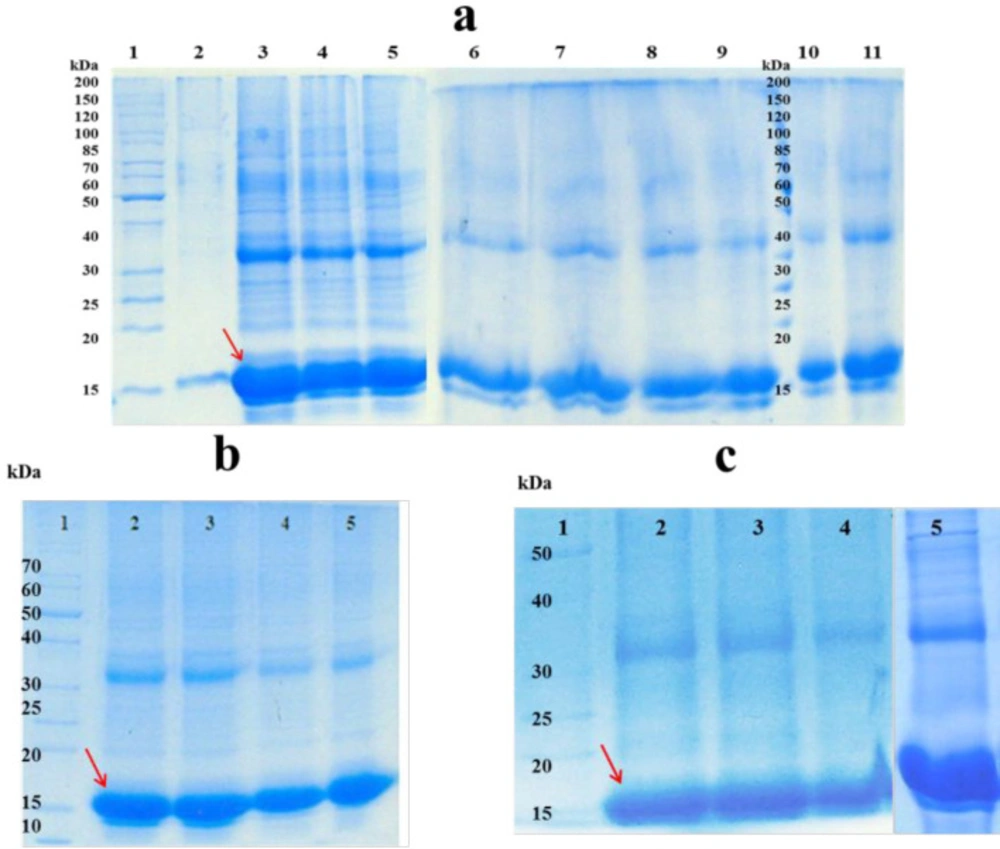
Soluble fractions after solublization of hGM-CSF inclusion bodies; a) effect of different concentrations of Urea and GdnHCl and their combinations on solubilizing inclusion bodies: Lane 1: protein marker (26614); Inclusion bodies were dissolved in 2, 4, 6 and 8 M Urea (Lanes 2-5). Inclusion bodies were dissolved in 2, 4, 6 and 8 M GdnHCl (Lanes 6-9). Inclusion bodies were dissolved in 2 M Urea + 2 M GdnHCl (Lane10), 4 M Urea + 4 M GdnHCI (Lane11).; b) Lane 1: protein marker (26614); Inclusion bodies were dissolved in 4 M Urea at pH = 11, 9, 7, 5 (lanes 2-5). c) Lane 1 protein marker (26614); Inclusion bodies were dissolved in urea 4 M at pH = 9 (lane 2); Inclusion bodies were dissolved in 4 M Urea at pH = 11 supplemented with different additives: 4 mM DTT (lane 3); 4 mM n-propanol (lane 4); 4 mM β-mercaptoethanol (lane 5).
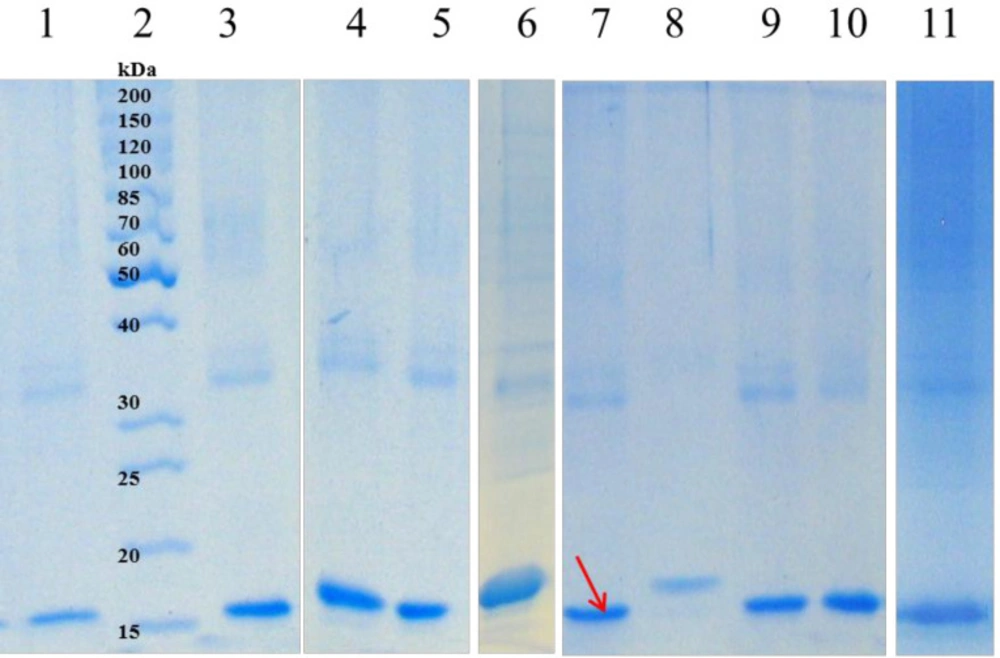
Soluble fractions after refolding of solubilized hGM-CSF inclusion bodies; Lane 2: protein marker (26614); solubilized inclusion bodies were diluted in refolding buffer containing: water (lane 1); 4 M Urea, 4mM β-mercaptoethanol, pH = 9 (lane 3); Citric acid (lane 4); Tricine (lane 5); Triton X-100 (lane 6); Imidazole (lane 7); Sorbitol (lane 8); SDS (lane 9); Urea (lane 10) or Ethanol (lane 11).
Normal (%) probability plot of the ‘studentized’ residuals for the model of hGM-CSF refolding
(A) Response surface plot for refolding of hGM-CSF shows the interaction between two factors in concentration of refolded hGM-CSF (µg/mL) by keeping other factor constant. (B) Large scale refolding of hGM-CSF with the optimum buffer; Solubilized hGM-CSF inclusion bodies were diluted in the optimum buffer additive condition (Lane 1) and protein marker (Lane 2)
Purification of hGM-CSF (A) Gel filtration (Superdex 200) chromatogram of hGM-CSF. The arrow indicates the peak of eluted hGM-CSF. (B) The peak fractions of gel filtration (140-147 min) were analyzed by SDS-PAGE. The arrow indicates eluted hGM-CSF. (C) SDS-PAGE analysis of hGM-CSF purified using Ni-NTA column: Solubilized hGM-CSF before loading onto column (Lane 1); The fractions were folded and eluted under hybrid condition using the native elution buffer containing 100 (Lane 2), 200 (Lane 3) and 400 (Lane 4) mM imidazole
Circular dicroism spectra of refolded hGM-CSF compared with standard hGM-CSF
Biological assay of hGM-CSF using MTT assay (A) Effect of refolded hGM-CSF and standard hGM-CSF on viability of HL-60 cells (b) Effect of refolded hGM-CSF on viability of HL-60 and U937 cells. Cells were incubated for 48 h with different concentrations (1-100 pg./mL) of hGM-CSF. The vertical bars indicate the standard deviations (n = 4).