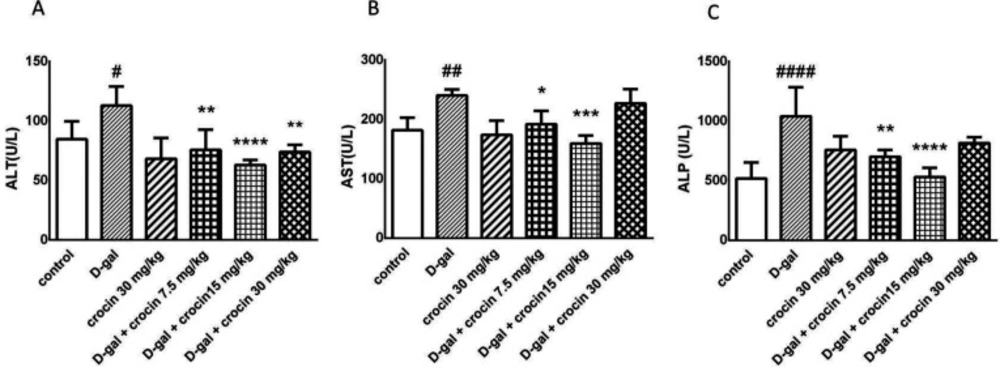
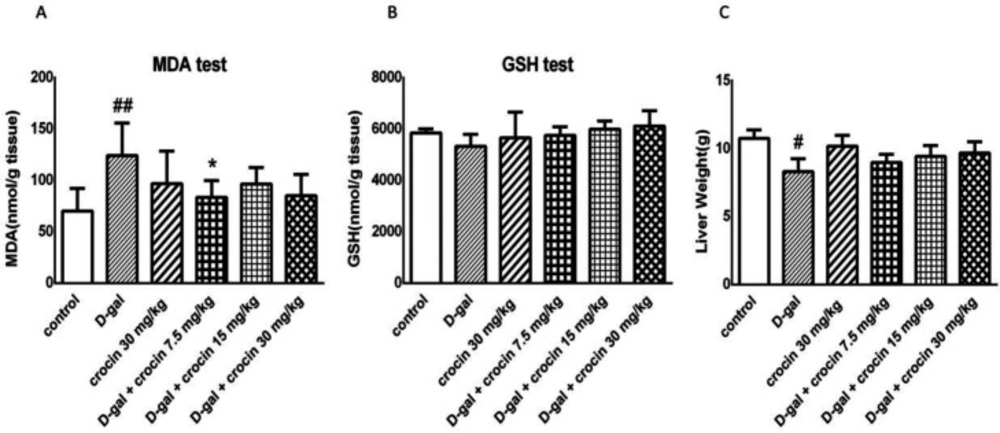
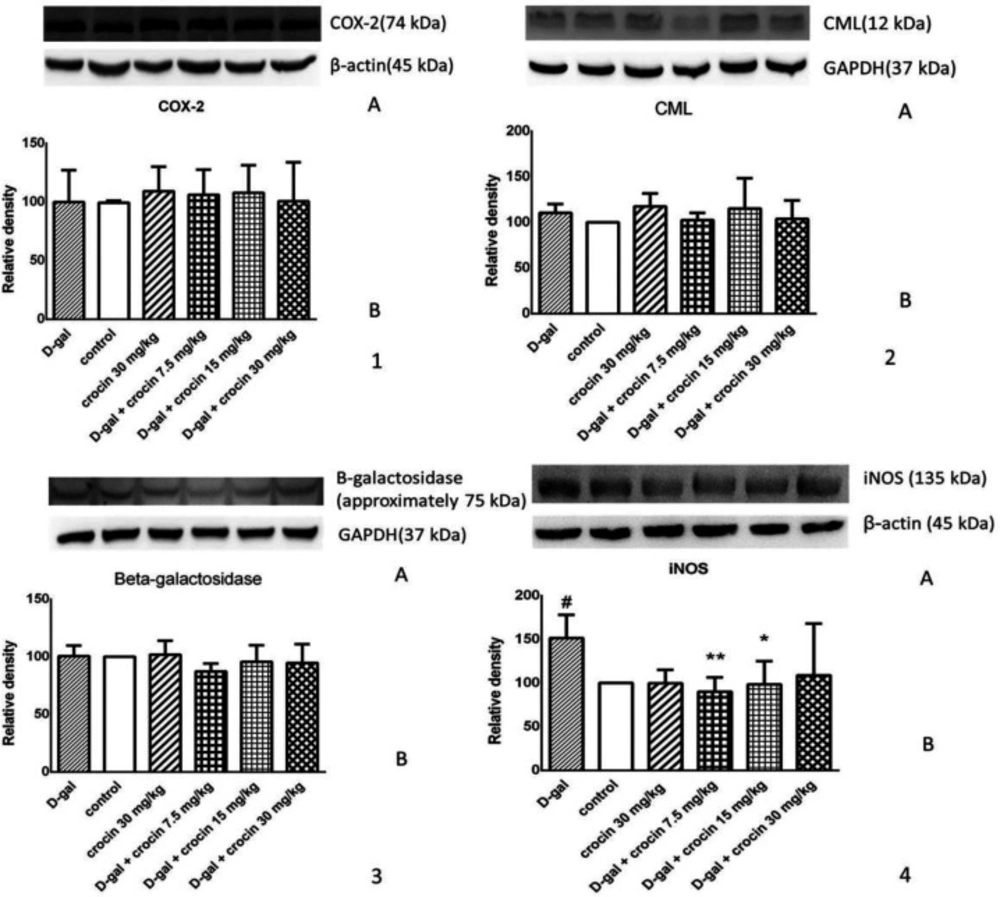
This study evaluated protective effects of crocin against D-gal-induced hepatotoxicity. Crocin prevented the liver damage that it is shown by ALT, AST, and ALP markers. The excessive release of these enzymes was significantly inhibited by crocin administration. Crocin suppressed the overgeneration of lipid peroxidation products such as MDA. iNOS was elevated by D-gal administration and was returned to the normal extent by crocin.
The most commonly mentioned mechanism for the development of the aging model by chronic D-gal administration is oxidative stress, ROS accumulation, and subsequent destructive events (
11-16,
36). Taken together, we believe that crocin as a powerful antioxidant and radical scavenger (
21), reversed all aging-like effects which were induced by D-gal administration in some biological aspects. Its evidence is decreased level of biochemical factors representing liver function such as ALT, AST, ALP, and also suppression of lipid peroxidation and MDA overproduction. Our findings demonstrate that in contrast to general expectation and some previous-mentioned experimental studies, GSH level did not have a significant change in the liver due to D-gal oversupplying. Hadzi-petrushev
et al. (2015) assessed the alterations of antioxidant enzymes induced by D-gal in different ages in liver and kidney of rats. They suggested that hepatic catalase (CAT) and glutathione peroxidase (GPx) activity, were even raised in all age groups of rats and glutathione reductase (GR) activity possessed approximately no difference between control and D-gal group in elderly rats. The increment of antioxidant activity is known as an adaptive response to neutralizing oxidant agents (
37). The findings, obtained from the study done by Samarghandian
et al. (2016) demonstrated that CAT enzyme level in kidney of the rats with 10 months old did not change and crocin was not able to increase CAT, SOD, and GPx significantly in these animals but the alterations in rats with 20 months old was considerable (
38). On the other hand, in Kasapoglu and Ozben study (2001), dispensability of aging-related reduction in antioxidant capacity was established (
39). In our study, enzymatic antioxidants rather than GSH may have changed due to D-gal administration.
There is a body of convincing evidence that oxidative stress and free radicals play an essential role in obesity promotion, insulin resistance, and subsequently diabetes (
18). Oh
et al. (2016) showed that in aged mice, insulin sensitivity decreased and glucose-dependent insulin secretion increased through overexpression of Ca sensing receptors. It should be considered that in this study, non-fasting blood glucose level did not change with age in 4, 8, 12 and 20 months old mice. These findings give us insight into the fact that aging does not always lead to hyperglycemic status (
17). Streptozocin as a diabetogenic agent acts via induction of oxidative stress and reduction of brain-derived neurotrophic factor (BDNF) in pancreas, brain, and liver (
40). In an experimental study crocin had protective effects on insulin resistance and hyperlipidemia induced by Streptozocin in the rats (
41). These approaches made us curious about D-gal that whether it has effects on blood glucose and insulin (through ROS generation) and also the effects of crocin on these likely metabolic changes. There are some
in-vivo studies in which galactose was used to generate diabetic cataract model in animals such as dogs or rats. However, galactose dosage and duration of its administration was more than our study (
42-
44). In our experiment, with a 56-day injection of D-gal (400 mg/kg/day), no insulin resistance was observed. Similar to our study, song
et al. (1999) indicated that D-gal (50 mg/kg/day SC) failed to cause diabetes model in mice, probably because of its different mechanisms responsible for aging simulation rather than diabetes induction (
16).
As our data represent, iNOS overexpression was inhibited by crocin administration. Nitric oxide can participate in a reaction with superoxide to generate reactive oxygen and nitrogen species. indeed, the most important point to drive ROS production and aging process is the NO/superoxide imbalance. It has also been exhibited that NO have reversible hampering effects on cytochrome c oxidase of mitochondria, that subsequently leads to less O
2 consumption and ROS formation. Based on this inference, some researchers believe that decreased NO level is a trigger for aging, thus reduction in NO level will be followed by increment of O
2 consumption and ROS production (
3,
45). Our findings support some investigations that have the same results as ours (
4,
26,
46).
It was reported that β-galactosidase is one of the most popular biomarkers for detection of senescent cells (
2). Basically, senescence associated β-galactosidase (SA-β-gal) activity is measured by staining of the cells or tissues with a chromogenic agent, but its level can be determined via western blotting as well, although its alterations are less observable by this method. The study was conducted by Lee
et al. (2006) demonstrated that the origin of SA-β-gal is the cell lysosome and the variations in SA-β-gal is a result of the changes in the number or activity of these organelles. The elevation of the β-galactosidase level is due to the aging process and the stop in cell replication, not the reason of that. Therefore, after discontinuity of the cell proliferation and accumulation of inactive or dead cells in tissue, the amounts of lysosomes and their enzymes will rise (
9). In regard to recent explanation, we can conclude that if aging process induces apoptosis in cells, the β-galactosidase may even decrease its level. Furthermore, it has been shown that the long term D-gal administration can result in the β-galactosidase rise but with some different methods from that of our investigation. In Ruan
et al. study (2013), the method of detection was tissue staining (
15). In the other study, which was carried out by Huang
et al. (2013), aging model was induced by D-gal intraperitoneal injection for 12 weeks (150 mg/kg/day) and the time of exposure was longer than that of our study (
36). Our treatment (with higher dose and 4 weeks shorter duration of D-gal injection) might cause more cell apoptosis and so, had no significant changes in β-galactosidase level.This hypothesis is also supported by liver weights as well. The decrement of liver weights in the D-gal group is possibly due to tissue necrosis and cell apoptosis as shown in some histomorphological assessments (
14).
Another complication in the aging phenomenon is the generation of advanced glycation end-products (AGEs) such as CML that can lead to the various harmful outcomes, for example predisposing to diabetes, neurodegenerative disorders, rheumatoid arthritis (
10), cardiovascular events, and arterial stiffness. This CML rise is significantly higher in elderly human than the old rats, because of their faster protein turnover and shorter lifetime (
8). The excessive production of CML during D-gal-induced aging has been demonstrated in cell culture and animal models (
16,
47). In our experiment, similar results to previous studies were obtained, but were not significant.
Cyclooxygenase-2 (COX-2) is mostly an inducible isoform of COX enzyme in the inflammatory condition, which is responsible for the synthesis of prostaglandins (PGs) from arachidonic acid. These molecules play an imperative role in both physiological and pathological pathways. A systematic study was performed by Kirkby
et al. (2016) showed that constitutive COX-2 also exists, with the greatest level of expression in the uro-renal, gastrointestinal and central nervous systems (
48). Oxidative stress can connect to the inflammation and immune system via Toll-like receptors (
6) and lipid peroxidation is able to activate the components of immune system, too (
7). The most common regulatory agent for COX-2 expression is NF-κB transcriptional factor (
48). As it is reported by Poligone and Baldwin (2001), NF-κB activity increases via several signaling factors and induces COX-2 expression. This enzyme produces PGE
2 in the early stage of inflammation and PGJ
2 and PGA in the late stage of inflammation. Therefore, at first, PGE
2 leads to more NF-κB level and subsequent events such as IL-8 expression, but finally, PGJ
2 and PGA enhancement creates a negative feedback on NF-κB activity and causes COX-2 suppression (
49). In some previous studies, COX-2 level was assessed after induction of aging model by D-gal in mouse kidney (
46), mouse liver (
50), rat brain (
51), and rat prostate (
52) and their findings exhibited that COX-2 level elevated in comparison with the control group. Yan
et al. (2016) simulated acute aging model in rat liver by intraperitoneal injection of 20 μg/kg of LPS and after 15 min, 700 mg/kg of D-galactosamine. They observed that COX-2 level in the aging-model group increased (
53). In our study, surprisingly, D-gal administration (400 mg/kg/day, for 56 days, SC) did not significantly change the COX-2 level in rat liver.
In conclusion, decreased level of ALT, AST, and ALP level by crocin in D-gal-treated rats revealed that this valuable compound has protective effects on hepatocytes. Crocin as a powerful antioxidant and radical scavenger was able to decrease MDA, a major product of lipid peroxidation, and inhibit iNOS and subsequently RNS overproduction which is associated with the aging process. Therefore, it can be considered for therapeutic purposes or as a useful supplement.
Effects of D-gal and crocin administration on ALT (A), AST (B) and ALP (C) level. The data are displayed as mean ± SD (n = 4). #P < 0.05, ##P < 0.01 and ####P < 0.0001 vs. control group; *P < 0.05, **P < 0.01,***P < 0.001 and ****P < 0.0001 vs. D-gal-treated group. D-gal: D-galactose
Effects of D-gal and crocin administration on Insulin (A) and blood sugar (B) level. The data are expressed as Mean ± SD (n = 5). D-gal: D-galactose
Effects of D-gal and crocin administration on MDA (A), GSH (B) levels and liver weight (C).The data are expressed as Mean ± SD (n = 4). ##P < 0.01 vs. control group; *P < 0.05 vs. D-gal-treated group. D-gal: D-galactose
Effect of crocin and D-gal on the protein levels of (1) COX-2, (2) CML,and (3) β-galactosidase. (1A), (1B) and (1C) are western blot images of COX-2, CML and β-galactosidase, respectively. (2A), (2B) and (2C) are determined using densitometric analysis. The data are presented as Mean±SD (n = 4). D-gal: D-galactose. (4) Effect of crocin and D-gal on the protein level of iNOS. (4A) is western blot images of iNOS and β-actin. (4B) is determined using densitometric analysis. The data are presented as Mean ± SD. n = 4. #P < 0.05 vs. control group; *P < 0.05, **P < 0.01 vs. D-gal-treated group