
Introduction
Experimental
Results
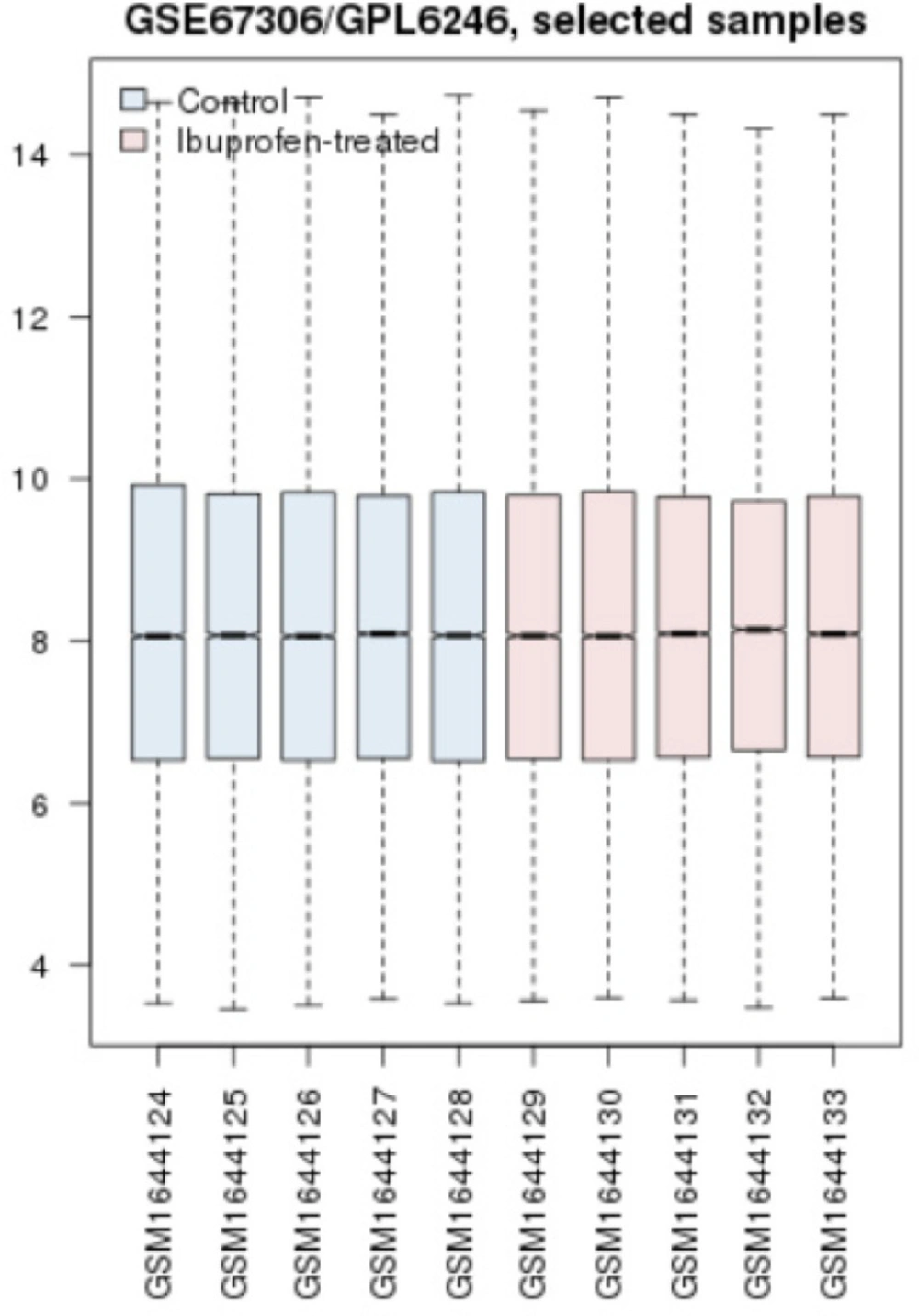
Cross comparison of two groups of AD mice and AD mice treaded with ibuprofen. The blue boxes refer to controls and the pink ones refer to AD-treated samples
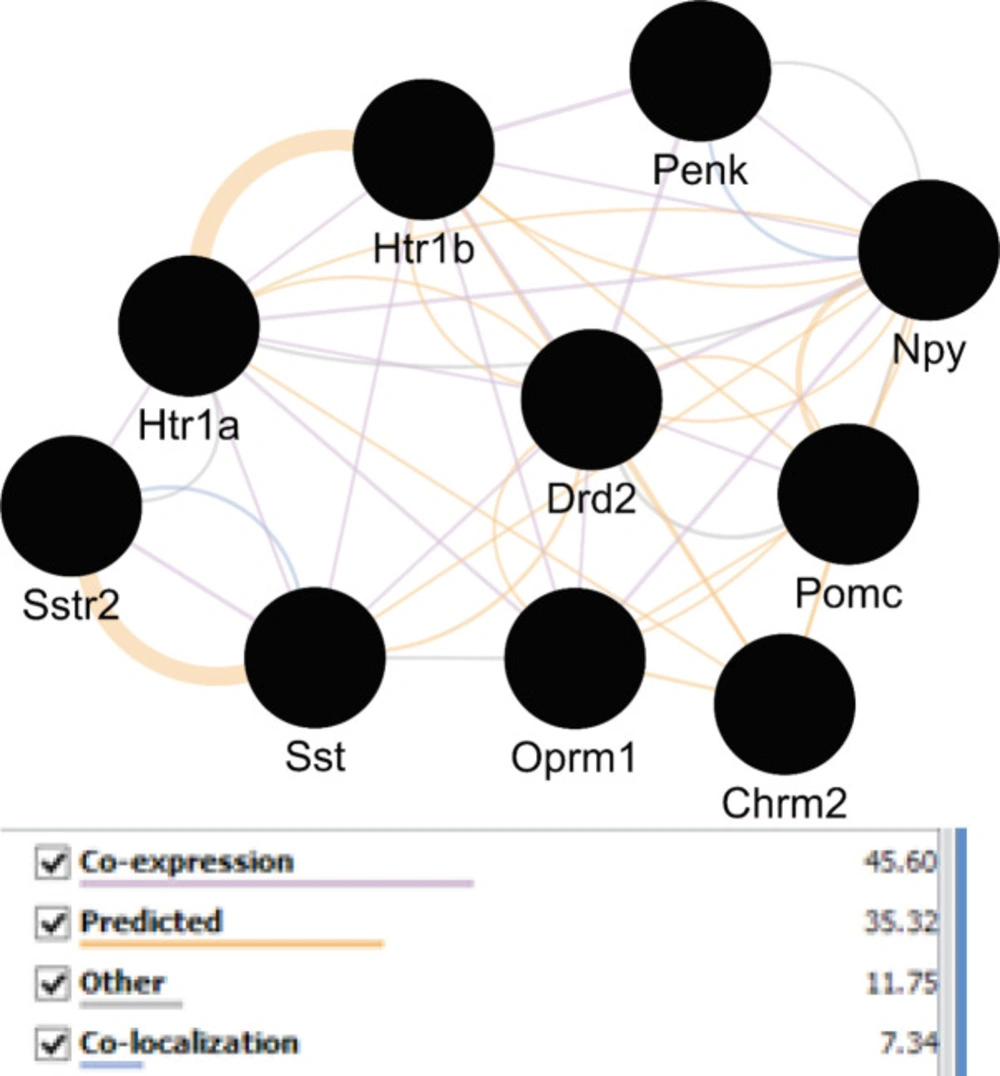
Network view of central elements of the main network corresponding to each other via different relationships. Different edges colors implies on different associations between these nodes
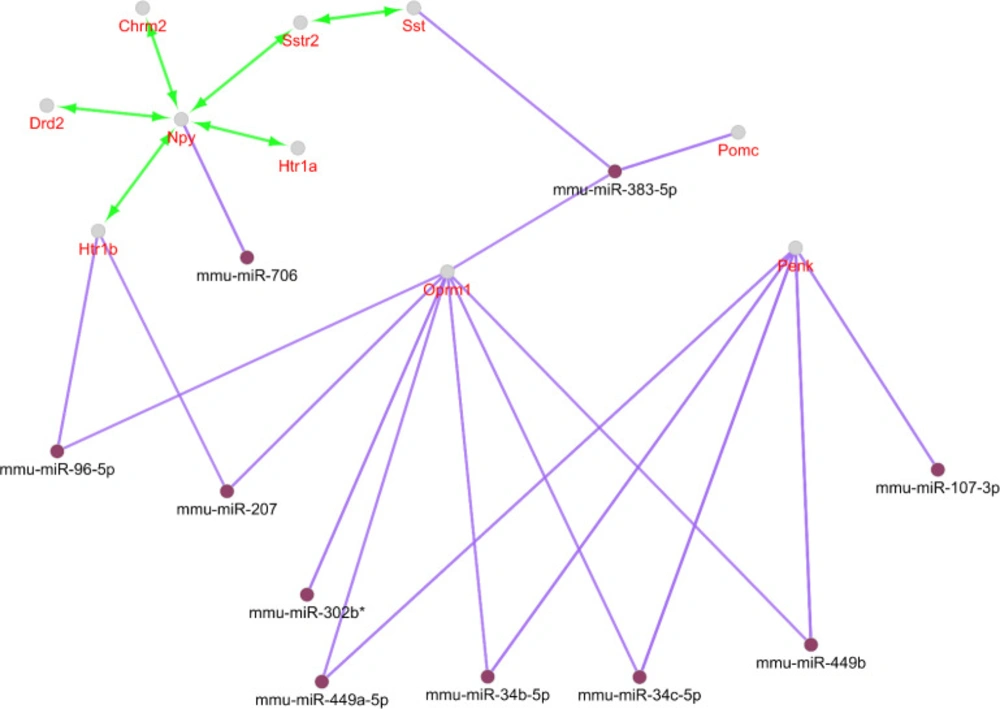
Action view and regulatory network for the central nodes of the original network. The score cut off for miRNA interactions is set to 0.6. A cut off 10 miRNAs are set for target central nodes. The actions include activation (green), expression (yellow), and inhibition (red). However, the last two are not concluded in any result. The edge thickness indicates the max and min scores (cut off for these action scores was set to 0.5 (medium)). * refers to the maximum kappa score value (0.796).

A sub-network including PENK and 10 relevant genes. The nodes are layout based on degree value. Bigger size and blue color refer to more connections. STRING application of Cytoscape is used to construct the sub network
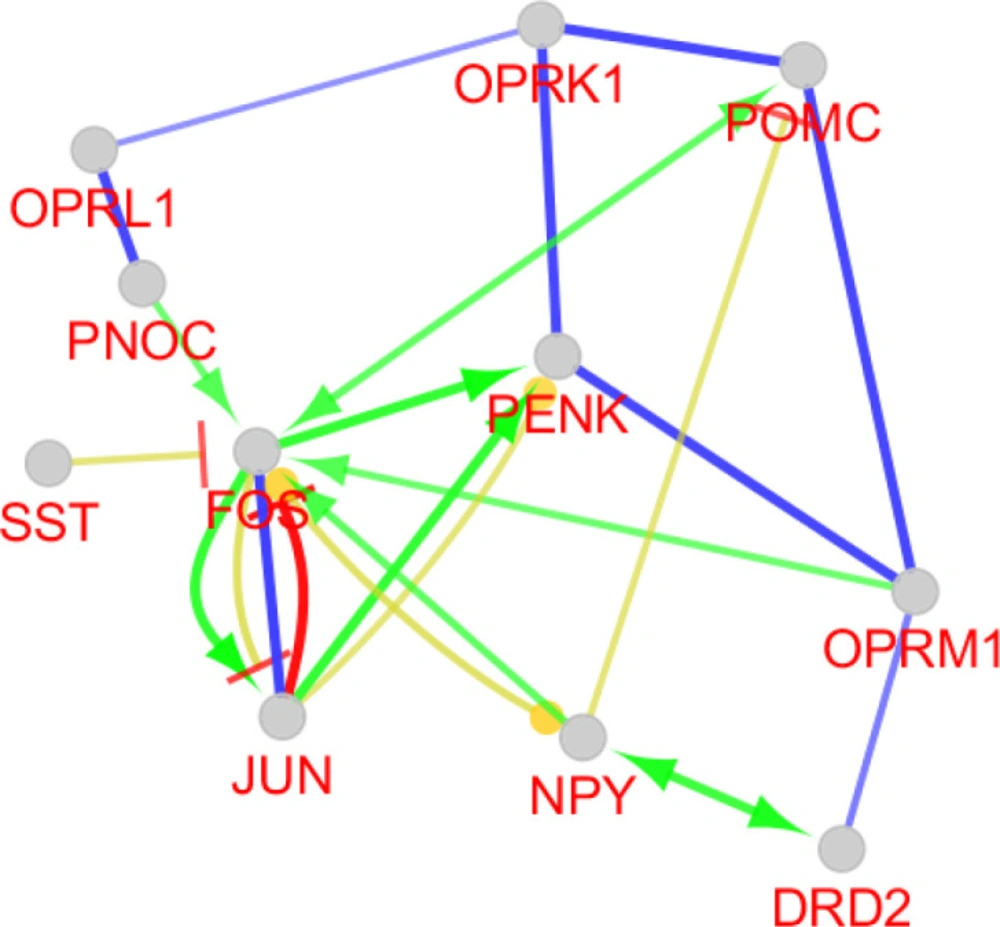
Action map including activation (green), inhibition (red) expression (yellow), and binding (blue) for PENK and its 10 relevant genes is presented. CluePedia application of Cytoscape is used to construct the action map
| GEO SAN | Samples |
|---|---|
| GSM1644124 | Hippocampus_APPS_Con_rep1 |
| GSM1644125 | Hippocampus_APPS_Con_rep2 |
| GSM1644126 | Hippocampus_APPS_Con_rep3 |
| GSM1644127 | Hippocampus_APPS_Con_rep4 |
| GSM1644128 | Hippocampus_APPS_Con_rep5 |
| GSM1644129 | Hippocampus_APPS_Ibu_rep1 |
| GSM1644130 | Hippocampus_APPS_Ibu_rep2 |
| GSM1644131 | Hippocampus_APPS_Ibu_rep3 |
| GSM1644132 | Hippocampus_APPS_Ibu_rep4 |
| GSM1644133 | Hippocampus_APPS_Ibu_rep5 |
| Row | Gene Name | FC | P-value | Regulation |
|---|---|---|---|---|
| 1 | Penk | 1.7 | ≤ 0.0007 | Up |
| 2 | Dsp | 1.9 | ≤ 0.0010 | Down |
| 3 | Ecel1 | 1.6 | ≤ 0.0010 | Up |
| 4 | Il1rl | 1.5 | ≤ 0.0020 | Down |
| 5 | Tdo2 | 2.5 | ≤ 0.0030 | Down |
| 6 | zcchc5 | 1.6 | ≤ 0.0040 | Down |
| 7 | Gm7120 | 1.7 | ≤ 0.0070 | Down |
| 8 | snord115 | 1.4 | ≤ 0.0100 | Up |
| 9 | Eif2s3y | 1.4 | ≤ 0.0300 | Down |
| Row | Name | Description | K | BC | CC |
|---|---|---|---|---|---|
| 1 | Htr1a* | 5-hydroxytryptamine (serotonin) receptor 1A | 72 | 0.23 | 0.62 |
| 2 | Sstr2* | somatostatin receptor 2 | 70 | 0.10 | 0.59 |
| 3 | Drd2* | dopamine receptor D2 | 70 | 0.13 | 0.61 |
| 4 | Htr1b* | 5-hydroxytryptamine (serotonin) receptor 1B | 70 | 0.10 | 0.60 |
| 5 | Penk | Preproenkephalin | 69 | 0.00 | 0.59 |
| 6 | Pomc | pro-opiomelanocortin-alpha | 69 | 0.00 | 0.59 |
| 7 | Oprm1 | opioid receptor, mu 1 | 69 | 0.00 | 0.59 |
| 8 | Npy | neuropeptide Y | 69 | 0.00 | 0.59 |
| 9 | Sst | Somatostatin | 69 | 0.00 | 0.59 |
| 10 | Chrm2* | cholinergic receptor, muscarinic 2 | 68 | 1.00 | 0.66 |
