Chemical Composition of C. botrys Essential Oil
The essential oil of C. botrys was isolated from the dried aerial parts of the plant through a hydro-distillation process with a yield of 0.97% (w/w) based on the dry weight of C. botrys. In total, 37 constituents were detected in the herbal essential oil by GC-MS analysis, as well as their composition and retention index. Accordingly, the essential oil of C. botrys was composed of monoterpenes (0.92%), oxygenated monoterpenes (2.68%), sesquiterpenes (13.71%), oxygenated sesquiterpenes (64.17%), and other components (4.06%). The major constituents of C. botrys essential oil were α-Eudesmol (16.81%), Elemol acetate (13.2%), Elemol (9.0%), and α-Chenopodiol-6-acetate (7.9%).
Cytotoxicity Induction by C. botrys Essential Oil
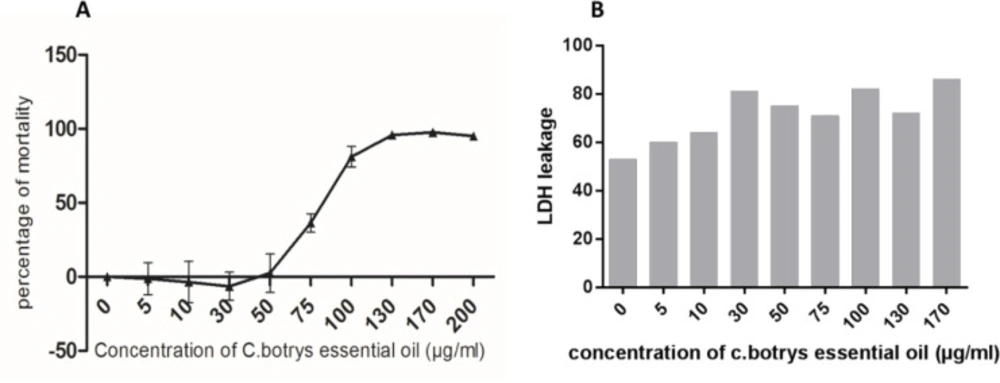
In the present study, a broad spectrum of the cytotoxic effects
C. botrys essential oil on HeLa human cancer cell line was investigated using the MTT assay. Cell viability upon the treatment of cells with increasing concentrations of the herbal essential oil (5-200 µg/mL) was assessed for 24 h. According to the results, the essential oil of
C. botrys exhibited concentration-dependent cytotoxic effects on the cell line at 24 h. Furthermore,
C. botrys essential oil exerted significant cytotoxic effects on the HeLa cells with the IC50 of 79.62 µg/mL after treatment for 24 h (
Figure 1).
Apoptosis Induction by C. botrys Essential Oil in Cervical Cancer Cells
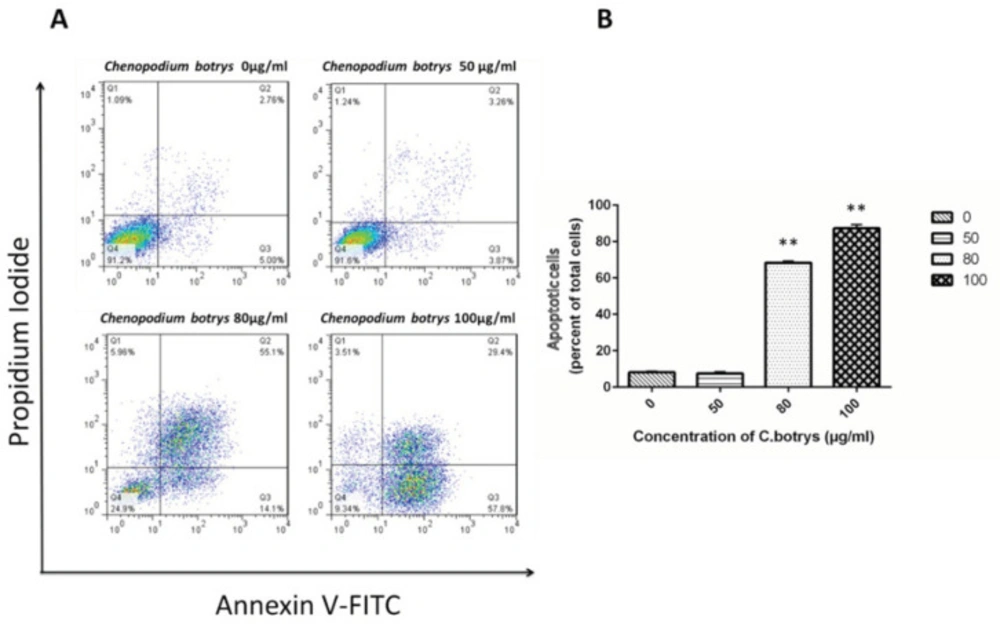
To determine whether the loss of the cell viability of cervical cancer cells induced by C. botrys essential oil was associated with the induction of apoptosis, the HeLa cells were treated by C. botrys essential oil, and the number of the apoptotic cells was determined using the Annexin V-FITC and PI detection kit as previously described.
Apoptotic cells were characterized as early (Annexin V+) and late (Annexin V+/PI+) apoptotic cells. Treatment of the HeLa cells with the essential oil of
C. botrys led to a significant dose-dependent increase (
p ˂ 0.001) in the number of apoptotic cells from 0 µg/mL to 80 µg/mL and 100 µg/mL, as follows: 0 µg/mL (mean ± SEM: 8.14±0.65), 50 µg/mL (7.53 ± 0.93), 80 µg/mL (68.4 ± 0.91), and 100 µg/mL (87.3 ± 1.95) (
Figure 2).
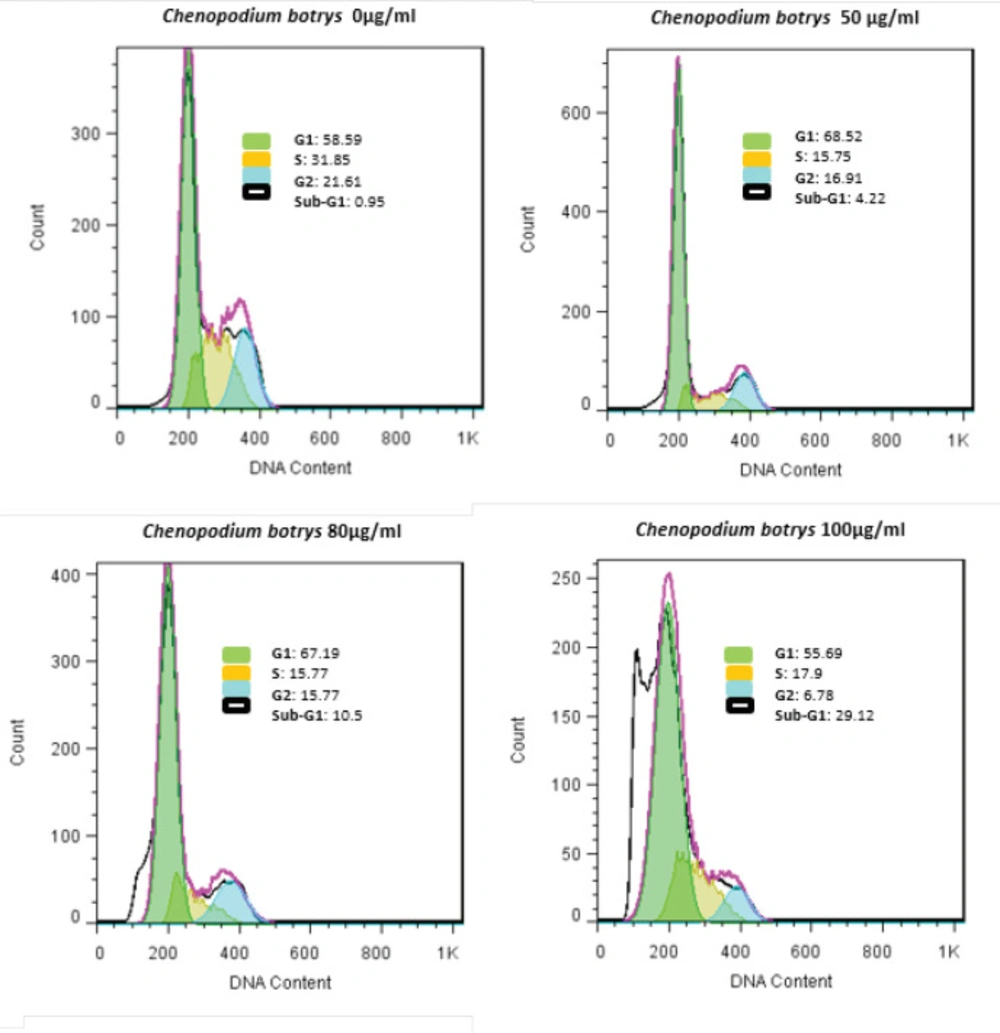
Induction of Cell Cycle Arrest in G1 Phase in Cervical Cancer Cells by C. botrys Essential Oil
Percentage of the cervical cancer cells in various phases of the cell cycle was investigated using the flow cytometry of PI-labeled HeLa cells after 24 h of treatment with
C. botrys essential oil. Treatment of the HeLa cells by
C. botrys essential oil caused an increase in the G0/G1 phases and a reduction in the G2 and S phases, which indicated cell cycle arrest in the G1 phase (
Figure 3).
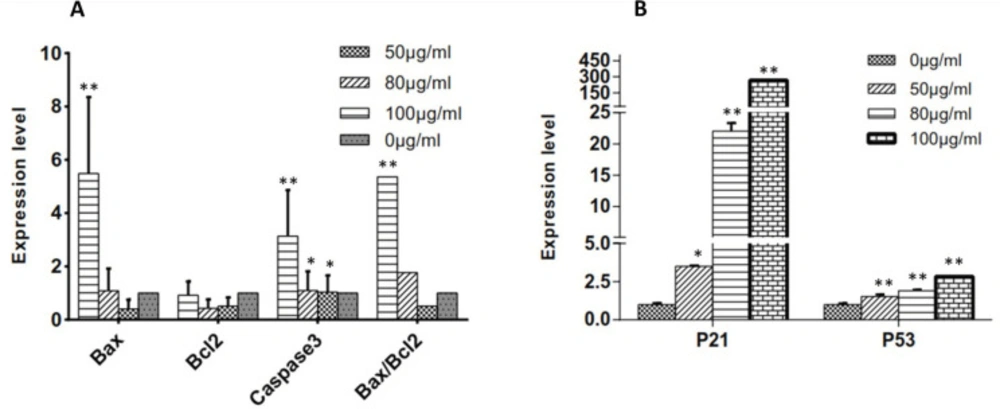
Effects of C. botrys on the Expression of Bcl-2 Family Members and Caspase-3
To evaluate the status of the intrinsic signaling of apoptosis in mitochondria, mRNA expression levels of the pro-apoptotic (
Bax) and anti-apoptotic genes (
Bcl2) were measured using real-time PCR. Moreover, we examined
caspase involved in
C. botrys-induced apoptosis by evaluating the mRNA expression level of
caspase-3. The essential oil of
C. botrys significantly increased the expression of
Bax (
p ˂ 0.001, between 0 µg/mL to100 µg/mL) and
caspase-3 (
p ˂ 0.001). Furthermore, the ratio of
Bax/
Bcl2 had a five-fold increase from the concentration of 50 to 100 µg/mL (
p ˂ 0.001).
Bcl2 gene expression had no significant difference between the groups (
Figure 4A).
Effects of C. botrys Essential Oil on the Expression of p21 and p53
To assess the status of the HeLa cells in cell growth arrest, we evaluated the expression levels of
p21 and
p53 genes. According to the findings, the essential oil of
C. botrys significantly increased the expression of
p21 (
p ˂ 0.001) and
p53 (
p ˂ 0.001) in a concentration-dependent manner, especially in the case of
p21 (
Figure 4B).
Phytochemicals may affect the signaling pathways within the cells, including those involved in the regulation of cell proliferation and activation of apoptosis(
20). Apoptosis, also known as programmed cell death, is a pathway through which cells undergo death to control cell proliferation or react to DNA damage (
21). Activation of these pathways is a major mechanism, through which cytotoxic drugs eradicate tumor cells (
22). In the current research, we evaluated the apoptosis induced by
C. botrys essential oil in HeLa cells and determined the involved molecular mechanisms that occur via the intrinsic pathway.
The results of GC/MS analysis showed 37 compounds representing 85.54% of the total contents of the essential oil (
Table 2). Moreover, the results indicated that
C. botrys is an abundant source of monoterpenes, especially sesquiterpenes (78.88%). Several studies have demonstrated that the major compounds of
C. botrys essential oil are α- and β-eudesmol(
23,
24), α- and β-chenopodiol (
25), camphor, and elemol (
26). However, the main components were noted as α-eudesmol (16.81%), Elemol acetate (13.2%), Elemol (9.0%), and α-Chenopodiol-6-acetate in the current research. In the present study, we investigated the cytotoxic effects of
C. botrys essential oil on cervical cancer cells, and the herbal essential oil was considered to be a potential cytotoxic agent in the treatment of HeLa cells by killing the cancer cells in a dose-dependent manner. In particular, treatment of the HeLa cells with 100 µg/mL of
C. botrys essential oil for 24 h could reduce the viability of cervical cancer cells by 81.14 ± 6.87% (IC50:79.64 µg/mL), which indicated the potential to eliminate cancer invasiveness.
The anti-tumor effects of the herbal essential oil could be attributed to cytostatic and/or cytotoxic function. Cytostatic substances are able to prevent cell growth and/or induce cell cycle arrest at various cell cycle checkpoints. On the other hand, cytotoxic substances induce cell death through apoptosis or necrosis (
29). Therefore, we demonstrated a mild elevation in the leakage of LDH by increasing the concentration of the essential oil. Moreover, LDH leakage had a significant adjustment at the IC50 dosage of
C. botrys essential oil.
In the current research, apoptosis was followed by cell cycle arrest in the G1 phase, which increased the cells in the sub-G1 (G0) phase and decreased cells in the G2 and S phases (
Figures 2 &
3). This finding is supported by the overexpression of
p21 and
p53 genes.
P21 is known to act as an essential inhibitor of CDK2, which is activated in response to various cellular and environmental signals to improve tumor suppressor events (
30).
In the present study, the wild-type tumor suppressor
p53 and
p21 of the HeLa cells was up-regulated (
Figure 4B). Therefore, it could be concluded that the up-regulation of
p53 by
C. botrys essential oil triggers the accumulation of
p21 protein, leading to G0/G1 phase cell cycle arrest and apoptosis in HeLa cells (
31-
33). It is also notable that
p53/
p21 is involved in the control of cell cycle, apoptosis, and maintenance of genomic stability (
34). Once DNA damage occurs, wild-type
p53 is expressed to induce cell cycle arrest at various checkpoints in order to repair the damaged DNA. As a result, unrepaired DNA cells are targeted by apoptosis (
32). Furthermore, this is accompanied by an increase in the expression of the
Bax and
Bax/
Bcl2 ratio. The cytosolic section of
p53 activates the apoptotic-effector protein
Bax to initiate apoptosis (
35). The damaged cells could be repaired to maintain the integrity of the cell or be cleared by apoptosis to remove a cancerous cell through G1 phase arrest. Apoptosis is an aspect of mammalian cell behavior, which is of utmost importance in the growth and development in tumor oncogenesis.
Bcl2 and
Bax are the critical regulators of cell apoptosis with remarkable anti-apoptotic and pro-apoptotic effects, respectively (
36). According to the literature, susceptibility to apoptosis is likely to be determined based on the
Bax/
Bcl2 ratio as the key regulators of apoptosis located mainly at the outer membrane of the mitochondria and endoplasmic reticulum. In the present study, their regulation of apoptosis was mainly achieved through the mitochondria. Overexpression of
Bcl2 leads to resistance to apoptosis, whereas overexpression of
Bax results in increased apoptosis (
36,
37).
Our findings indicated the adjustment of the expression of Bcl2 and Bax compared to the control group. Bcl2 expression decreased significantly, while Bax expression had a statistically significant increase after treatment with C. botrys essential oil. Moreover, the Bax/Bcl2 ratio and caspase-3 elevated in the HeLa cells after treatment with C. botrys essential oil, which is consistent with the results regarding cell apoptosis.
According to the results of the present study, C. botrys essential oil induced intrinsic apoptosis through adjusting the expression of Bcl2 and Bax to enhance their ratio, followed by the triggering of the caspase cascade. Therefore, it could be concluded that C. botrys essential oil could potentially eliminate cancer cells by conducting apoptosis pathways.
A) Cytotoxic Effects of C. botrys on HeLa Cells Measured by MTT Assay; B) LDH Leakage Measured in Supernatant of Treated and Control HeLa Cells
HeLa Cells Treated with C. botrys Essential Oil (50, 80, and 100 µg/mL) for 24 H Analyzed by Flow Cytometry for Apoptosis; A) Lower right quadrant shows Annexin-V positive cells (early apoptotic), upper left quadrant shows PI positive cells (necrotic cells), upper right quadrant shows Annexin-V and PI positive cells (late apoptotic cells), and lower left quadrant shows PI negative and Annexin-V negative; B) Percentage of total apoptosis for each condition is characterized in a histogram. The number of apoptotic cells were significantly increased in 80 µg/mL (p ˂ 0.001) and 100 µg/mL (p ˂ 0.001) comparing to control group
HeLa Cells Treated with C. botrys Essential Oil (50, 80, and 100 µg/mL) for 24 H Analyzed by Flow Cytometry for Cell Cycle Assay
(A) Expression Levels of Bcl2 Family and Caspase-3 and Bax/Bcl2 Ratio at Various Concentrations of C. botrys Essential Oil (50, 80, and 100 µg/mL); (B) Expression Levels of p21 and p53 at Various Concentrations of C. botrys Essential Oil (50, 80, and 100 µg/mL). Comparison were done between groups. (**) was considered as pvalue ˂0.001 and (*) was considered for pvalue ˂0.05
| Gene | | Sequence | bp |
|---|
| Bax | Forward | CGCCCTTTTCTACTTTGCCA | 166 |
| Reverse | GTGAGGAGGCTTGAGGAGTC |
| Bcl2 | Forward | GCCTTCTTTGAGTTCGGTGG | 192 |
| Reverse | GAAATCAAACAGAGGCCGCA |
| Caspase-3 | Forward | ACTGGACTGTGGCATTCAGA | 162 |
| Reverse | GCACAAAGCGACTGGATGAA |
| p53 | Forward | CCCCTCCTGGCCCCTGTCATCTTC | 265 |
| Reverse | GCAGCGCCTCACAACCTCCGTCAT |
| p21 | Forward | CGCATGGGTTCTGACGGACATCC | 257 |
| Reverse | TGCCGAAGTCAGTTCCTTGTGG |
| GAPDH | Forward | GGAAGGTGAAGGTCGGAGTCA | 101 |
| Reverse | GTCATTGATGGCAACAATATCCACT |
| Class | RIL3 | RIL2 | RIL1 | Kovats Retention Index (RIa) | % | Rt (min) | Compounds | No. |
|---|
| MH | 991 | 990 | 981 | 981.8085 | 0.60 | 4.986 | β-Myrcene | 1 |
| MH | 1031 | 1030 | 1014 | 1017.147 | 0.32 | 6.143 | Limonene | 2 |
| MO | | | 1096 | 1073.13 | 0.97 | 8.239 | α-Thujone | 3 |
| MO | | 1145 | 1124 | 1126.591 | 0.22 | 10.377 | p-Menth-2-en-1-ol | 4 |
| MO | 1177 | 1180 | 1169 | 1162.908 | 0.35 | 11.923 | 4-Terpineol | 5 |
| MO | | 1193 | 1187 | 1177.402 | 0.52 | 12.54 | α-Terpineol | 6 |
| MO | 1285 | | 1285 | 1270.805 | 0.36 | 16.582 | Bornyl acetate | 7 |
| MO | 1354 | 1354 | 1354 | 1336.388 | 0.26 | 19.607 | citronellyl acetate | 8 |
| SH | 1391 | 1395 | 1382 | 1378.25 | 2.64 | 21.626 | β-elemene | 9 |
| SH | | 1425 | 1420 | 1398.155 | 1.42 | 22.586 | Trans-caryophyllene | 10 |
| SH | | 1460 | 1451 | 1433.083 | 0.28 | 23.955 | α-Humulene | 11 |
| SO | | | 1454 | 1442.879 | 0.23 | 24.334 | Trans-geranylacetone | 12 |
| SH | | | 1470 | 1465.96 | 0.52 | 25.227 | β-Eudesmene | 13 |
| SH | | | 1485 | 1484.053 | 1.13 | 25.927 | Aromadendrene | 14 |
| SH | | | 1505 | 1494.908 | 1.40 | 26.347 | γ-Cadinene | 15 |
| SH | 1524 | | 1516 | 1505.52 | 0.45 | 26.726 | δ-Cadinene | 16 |
| SO | | | 1539 | 1517.956 | 0.68 | 27.136 | α-Copaen-11-ol | 17 |
| SO | 1540 | | 1547 | 1537.307 | 9.02 | 27.774 | Elemol | 18 |
| SH | 1556 | | 1562 | 1551.319 | 0.85 | 28.236 | Germacrene B | 19 |
| SO | | 1583 | 1578 | 1561.541 | 0.95 | 28.573 | Caryophyllene Oxide | 20 |
| SO | | 1644 | | 1624.721 | 0.27 | 30.571 | γ-Eudesmol | 21 |
| SO | | 1653 | | 1641.754 | 16.81 | 31.074 | α-Eudesmol | 22 |
| SO | | | 1675 | 1667.931 | 13.17 | 31.847 | Elemol acetate | 23 |
| SO | | 1678 | | 1673.383 | 1.40 | 32.008 | botrydiol | 24 |
| SO | | 1691 | 1682 | 1678.835 | 1.13 | 32.169 | juniper camphor | 25 |
| SO | | | 1729 | 1696.749 | 0.42 | 32.698 | α-Costol | 26 |
| SO | | 1724 | 1736 | 1701.249 | 0.55 | 32.828 | guaiol acetate | 27 |
| SO | 1778 | | 1782 | 1770.242 | 5.05 | 34.706 | γ-eudesmol acetate | 28 |
| SO | 1789 | | 1785 | 1779.023 | 2.44 | 34.945 | α-eudesmol acetate | 29 |
| SO | | | 1791 | 1788.758 | 4.53 | 35.21 | β-Eudesmol acetate | 30 |
| SO | | 1830 | | 1866.967 | 4.30 | 37.223 | β-Chenopodiol | 31 |
| SO | | 1876 | | 1875.52 | 0.38 | 37.441 | α-Chenopodiol | 32 |
| SO | | 1904 | | 1883.837 | 0.55 | 37.653 | β-Chenopodiol-6-acetate | 33 |
| NH | | | 1928 | 1917.371 | 0.66 | 38.484 | Methyl Palmitate | 34 |
| SO | | 1957 | | 1930.473 | 2.15 | 38.8 | Acetoxyeudesman-4-α-ol-11 | 35 |
| SO | | 1977 | | 1940.796 | 7.88 | 39.049 | α-Chenopodiol-6-acetate | 36 |
| NH | | | 2128 | 2113.458 | 0.68 | 43.065 | Methyl Stearate | 37 |
| | | | | 85.54 | | Total Identified Chemicals | |
| | | | | 0.92 | | Monoterpenes (MH) (%) | |
| | | | | 2.68 | | Oxygenated Monoterpenes (MO) (%) | |
| | | | | 13.71 | | Sesquiterpenes (SH) (%) | |
| | | | | 64.17 | | Oxygenated Sesquiterpenes (SO) (%) | |
| | | | | 1.34 | | Non terpene hydrocarbon (NH) | |