This review aims to summarize different cellular mechanisms of Wnt signaling pathways and risks of diseases associated with dysregulated Wnt signaling pathways under the environmental exposure of heavy metals. A literature search was performed on different databases, including PubMed Central, Embase, Medline, and Google Scholar. Search terms were Wnt, canonical, noncanonical, signaling pathway, β-catenin, environment, and heavy metals used to sort the articles using Boolean operators. Published articles on Wnt signaling pathways were considered to summarize the cellular mechanism of canonical and noncanonical signaling pathways. At the same time, published articles on heavy metals as contributing factors for causing diseases via influencing Wnt signaling pathways were included to summarize environmental heavy metals’ effect via affecting the Wnt signaling pathway. Articles search remained limited to published articles in the English language only.
Environmental exposure of heavy metals and deregulated Wnt signaling pathways
Arsenic
Environmental arsenic (
As) exposure induces malignant transformation (
15-
17). Animal studies revealed that
As induces cancer cell survival, proliferation, and migration via modulating various signaling pathways such as
Wnt/β-catenin,
BMP7,
COX2, and influencing possible cross-talk among them (
18). Angiogenesis contributes to carcinogenesis. It promotes tumor growth, invasion, and metastasis via
β-catenin-VEGF pathway activation (
19-
21). A study on
As-transformed human bronchial epithelial cells demonstrated
As-induced an increase in vascular endothelial growth factor (
VEGF) expression, an angiogenic stimulating growth factor augments
β-catenin activity that leads to angiogenesis and risk of carcinogenesis (
20). A combination or single exposure of trivalent arsenic (
As(III)) or hexavalent chromium promoted colorectal tumor in azoxymethane/dextran sodium sulfate treated mice. As was found to induce tumorigenesis due to the
ROS-mediated
Wnt/β-catenin signaling pathway. As. by generating
ROS caused imbalance of oxidant and antioxidant enzymes along with declined superoxide dismutase (
SOD) and
catalase level, while increased expression of
β-catenin,
phospho-GSK,
NADPH oxidase1 (
NOX1), and
8-OHdG. Suggesting the role of
As. exposure to the tumor size increase, incidence, and inflammation via modulating the
Wnt/β-catenin signaling pathway (
22). Noncanonical
Wnts such as
Wnt5b are known to be associated with cancer and disease pathologies (
23,
24). Noncanonical
Wnt signaling regulates cell migration via activation of protein
kinase Cα (
PKCα) (
25). As exposure in the endothelial cells activates
Rac1,
i.e., required for remodeling and angiogenesis (
26). Persistent
As exposure upregulates
Rac1, Wnt5b, and
PKCα in
As-transformed cells suggesting the role of noncanonical
Wnt5b in
PKC activation, cell migration, and cancer risk (
27). Environmental
As exposure also promotes cancer via altering cell fate determination through
Wnt signaling pathway activation. In human mesenchymal stem cells,
As exposure upregulates the
Wnt3a protein and its
mRNA, while it inhibits the expression of
PPARγ, C/EBPα/β, and interaction between them, thus adversely affecting adipogenesis (
28). Since
PPARγ positively, while
Wnt negatively regulates adipogenesis (
28,
29). Moreover,
CCAAT enhancer-binding protein (
C/EBPs) expresses in adipocytes, whose inhibition impairs adipogenesis (
30,
31). Another study demonstrated changes in the adipose-derived mesenchymal stem/stromal cells (
ASCs) differentiation in mice vide
As induced altered canonical
TGFβ signaling pathway and dose-dependent decline in the
β-catenin (
CTNNB1), osteogenic such as
Runx2, OPN, and
BGP along with and chondrogenic such as
Sox9, DSPG3 and
ACAN gene expression (
32). As. exposure during embryogenesis of mice found to repress the muscle and neuron-related transcription factors, including
Pax3, Myf5, MyoD, myogenin, neurogenin 1 and
2, and
NeuroD. Such resulted in altered embryonic stem cells differentiation into skeletal muscles and neurons by repressing the
Wnt/β-catenin signaling (
33). Moreover, chronic
As exposure reported renal cancer via persistent decrease in
β-catenin expression
, declined
Wnt4, BMP7 and duration dependent increase in
Wilms’ tumor protein 1 (
Wt1),
Cox2, MMP2 and
MMP9 expression in
RIMM-18 cells (
18). The canonical
Wnt signaling pathway is vital to regulate nephron induction during the development of the kidney mediated by
Wnt4 (
34).
BMP7 promotes kidney repair after obstruction-induced renal injury (
35). Likewise,
Wt1 is essential for normal kidney development (
36). However,
matrix metalloproteinases and
Cox2 overexpression are associated with tumorigenesis (
37,
38). Suggesting
Wt-1,
Wnt4, and
BMP7 expression required in murine for normal kidney development.
As exposure induces renal cancer via affecting
BMP7, COX-2, and
Wnt/β
-catenin signaling pathways and possible cross-talk among them (
18). The facts above suggested that
As contributes to angiogenesis, carcinogenesis, and tumorigenesis via modulating directly or indirectly canonical and noncanonical
Wnt signaling pathways.
Cadmium
Agency for Toxic Substances and Disease Registry designated cadmium (
Cd) as a carcinogen due to its toxic effect via releasing
ROS, impairing calmodulin activity, and potential of altering signal transduction networks including
Wnt/β-catenin, and
estrogen (
39,
40).
Cd adversely affects immunity, leading to osteoporosis and bone diseases via modulating hematopoietic stem cells (
HSCs) and progenitor cells towards myelopoiesis (
41,
42).
Cdc42 is known to regulate
HSCs rejuvenation and aging via the noncanonical
Wnt5a signaling pathway (
43,
44). While
Cd exposure contributed toxicity to the immune system through impaired
HSC function and activated the noncanonical
Wnt5a-Cdc42 signaling pathway (
45).
Cd as an endocrine disruptor induces nuclear translocation of
β-catenin, causing increased expression of
Wnt/β-catenin target genes and
caspase3 activation in human osteoblastic Saos-2 cells. This resulted in osteoblastic apoptosis and necrosis due to altered bone homeostasis and the future risk of bone diseases (
46).
Wnt/β-catenin signaling is vital for vascularization and angiogenesis (
47). Environmental
Cd exposure increases the risk of cardiovascular diseases (
CVDs) via abnormal
Wnt/β-catenin signaling and aryl hydrocarbon receptor targets genes including
Ahr, Arnt, Nkx2.5, Ctnnb1 and
Gsk3β. Thus impairs the physiological function of
Ahr in regulating
Wnt/β-catenin signaling that leads to the risk of
CVDs (
48). Likewise, Japanese medaka embryos reported
Cd-induced adverse effects to the early life stages of fish via deregulated
Wnt signaling pathway. There observed negative impacts on heartbeat, cardiac morphogenesis, spinal and cardiac deformities and risk of
CVDs. There observed
Cd induced suppressed expression of DNA repair
rad51 gene, pro-apoptotic
bax gene, impaired mitochondrial respiration via inhibiting transcription of
NADH-dehydrogenase nd5 gene, and overexpression of cell proliferation and differentiation gene
i.e.
Wnt1 (
49).
Cd exposure also contributes to developmental defects among animals via modulating canonical and noncanonical
Wnt signaling pathways.
Cd induces varying degree of adherens junction breakdown in the periderm, disturbing
cadherins distribution and their intracellular associates via aberrant
Wnt signaling pathway resulted in ventral body wall (
VBW) defect (
50).
Rho-associated coiled-coil-containing protein kinase (
ROCK)
I and
ROCK-II regulates signaling from
Rho to the
actin cytoskeleton in
Wnt non-canonical signaling pathway while it absence demonstrated ventral body wall (
VBW) defect. A study on chick embryo demonstrated
Cd induced downregulated
ROCK I and
ROCK-II genes expression during embryogenesis that resulted into
VBW defect in chick embryo due disrupted
Wnt non canonical signaling pathway (
51). Noncanonical signaling pathways such as
Wnt/Ca2+ regulates cell movement and adhesion during embryogenesis.
Wnt is vital for
PKC activation and
calcium/calmodulin-dependent kinase II (
CaMKII) in the
Wnt/Ca2+ pathway requiring for actin-cytoskeleton organization and cell adhesion (
52,
53).
Cd treated chick embryos reported disrupt noncanonical
Wnt/Ca2+ signaling pathway via downregulation of
Wnt11, PKCα and
CaMK11 gene expression during embryogenesis, thus impairing cell movement and adhesion and risk of
VBW defects such as omphalocele (
54).
Cd reported carcinogenic activity via several mechanisms involving
Wnts.
Cd causes oncogenic transformation of normal cells by recruiting normal stem cells to an oncogenic phenotype by noncontagious carcinogen transformed epithelia via dysregulated
Wnt3 expression (
55). Thymocyte requires sonic hedgehog and
Wnt/β-catenin signaling pathways for its maturation. Environmental
Cd exposure in mice demonstrated decreased expression of these pathways in the thymus, thereby altering the expression of their target genes resulting in altered thymocyte development, increased cell proliferation and risk of cancer development (
56).
Cd exposure causes nuclear translocation of
β-catenin.
Cd also reduces the interaction between
β-catenin and
AJ components, including
α-catenin and
E-cadherin, thus increasing the binding of
β-catenin with
TCF4 transcription factor of
Wnt signaling pathway and thus upregulates
Wnt target genes including
Abcd1b, c-Myc and
cyclin D1. However,
E-cadherin overexpression reduces
Wnt signaling, cell proliferation and
Cd toxicity (
57). Chronic
Cd exposure via drinking water causes transcriptional activation of
Wnts and initiates epithelial to mesenchymal transition (
EMT), leading to renal fibrosis and the risk of developing cancer.
Cd exposure considerably increases kidney
Cd content which in turn increases expression of various
Wnt ligands, including -
3a,6,7a/b,9a/b,10a and
11 and upregulation of
Fz1 to
Fz10 except
Fz3 receptor. Thus caused increased expression of
Wnt target genes such as
Abcd1b, c-Myc and
cyclin D1 which promote cell proliferation, survival, migration and malignancy that leads to characteristic changes in the renal epithelial cells towards fibrosis and cancer through activated Wnt signaling pathway (
58). These facts suggesting that
Cd induces nephrocarcinogenesis via initiating
Wnt signaling pathway, disrupting
E-cadherin/β-catenin complex resulting in excessive nuclear translocation of
β-catenin and
TCF4 activation and upregulation of
MDR1, Abcd1b, c-Myc and
cyclin D1 genes (
59).
Chromium
Chronic exposure of hexavalent chromium (Cr) on BEAS-2B human lung epithelial cells demonstrated changes in the various gene expression mostly related to cell adhesion, protein ubiquitination, oxidative stress,
EMT, metastasis, and
Wnt signaling. There also observed upregulation of potential lung cancer biomarker ubiquitin carboxyl-terminal hydrolase L1 (
UCHL1) that initiates the transformation of lung epithelial cells towards an early stage of lung cancer (
60). Another study reported that chromium promoted colorectal cancer through
ROS-mediated
Wnt/β-catenin signaling pathway (
22).
Copper
Copper (
Cu) inhibits zebrafish egg hatching via suppressing embryonic motility (
61). It also impairs zebrafish swimbladder development and inflation by inhibiting the specification and formation of three swimbladder layers in a stage-specific manner (
62). These were due to
Cu-induced generation of
ROS and downregulation of
Wnt signaling (
61,
62). However,
Wnt agonist 6-bromoindirubin-3’-oxime (
BIO) was found to alleviate the suppressing effect of
Cu on egg hatching and swimbladder development (
61).
Cu induces toxicity to the early development of zebrafish (
63). Transcription factors such as
Ntl required for the development of posterior body structures (
64),
Dlx regulates intracellular signaling between neural and non-neural ectoderm and is vital for patterning adjacent cell fate (
65),
Hgg regulates the position of the anterior prechordal mesoderm (
66),
Wnt5 and
11 required for convergence and extension movement during various stages of gastrulation (
67).
Pax2 and
6 regulate
CNS development (
68), and cardiac
myosin light chain 2 (
Cmlc2) is an essential component of thick myofilament assembly while, its expression inhibits the cardiac looping resulting in impaired cardiac development (
63). Environmental
Cu exposure demonstrated toxicity to zebrafish by reducing the size of the head and eyes, aberrantly affect the dorsoventral patterning, cell migration of gastrulation, and prevent looping of heart tube during cardiogenesis. Such phenotypes were due to altered gene expression of
ntl, dlx3, and
hgg during gastrulation,
Cmlc2 expression, and decreased
pax2 and
pax6 gene expressions along with decreased
Wnt5 and
11 transcription factors (
63).
Lead
Environmental lead (
Pb) exposure
Pb induces neurotoxic and extra neurotoxic pathophysiological outcome that tends to sustain and maintain for a lifetime (
69). Developmental chronic
Pb exposure through lactation among rat pups demonstrated impaired learning and memory (
70). The role of
activity-regulated cytoskeleton-associated protein (
Arc/Arg3.1) and hippocampal
Wnt7a is known to regulate dendritic spines’ formation and structure (
71,
72). Dendritic spines are essential for excitatory synaptic transmission, and any change in their construction, numbers, and morphology will affect synaptic plasticity and spatial learning (
73). Chronic
Pb exposure reported the dose-dependent reduction of spine density and dentate gyrus region causing dysregulated synaptogenesis, impaired
Arc/Arg3.1 and hippocampal
Wnt7a ultimately resulted in impaired learning and memory among adult rats (
70). Several animal studies reported
Pb-induced bone pathologies such as osteoporosis, impaired healing of fractured bone, skeletal deficit growth, and development due to
Pb-induced modulation of the
Wnt/β-catenin signaling pathway and their related key regulators (
74,
75). It is well known that
Wnt/β-catenin signaling regulates osteoblastic anabolic function in bone formation (
76). Murine studies reported declined osteoblastogenesis due to
Pb exposure (
74,
75). This is due to
Pb-induced
sclerostin production via
TGFβ canonical signaling pathway (
74). Even low
Pb exposure increases
peroxisome proliferator-activated receptor-γ (
PPAR-γ) and
sclerostin while decreases
β-catenin and
Runx2 in stromal precursor cells, thereby disrupt bone homeostasis via inhibition of the
Wnt/β-catenin pathway (
75). Likewise, the subtoxic
Pb concentration was found to decrease
alkaline phosphatase (
ALP),
type 1 collagen (
COL1),
osteocalcin (
OC), and
Runx2 impairing regulation of
Wnt3a, Dkk-1, pGSK3β, and
β-catenin (
77). Environmental
Pb exposure also alters progenitor cell differentiation via promoting osteoclastogenesis and suppressing osteoblastogenesis, resulting in reduced trabecular bone quality, bone strength, and spine density due to reduced
Wnt signaling, thereby negatively impacting spine outgrowth (
78,
79).
Wnt signaling is also an important anabolic pathway required for chondrocyte maturation and endochondral ossification (
80). While
Pb is the potent inhibitor of endochondral ossification due to the deficit
Wnt/β-catenin signaling pathway that delays bone mineralization, causing the development of immature cartilage in the callus, thus impair healing of fractured bone (
81).
Pb induced upregulation of aggrecan,
Sox-9 and
type 2 collagen modulate multiple signaling pathways such as
AP-1, BMP, and
nuclear factor-kappa B (NF-kappaB) and
TGFβ, thus induce chondrogenesis (
82). Facts as mentioned earlier suggest that
Pb exposure via impairing the function of several key regulators of
Wnt/β-catenin signaling pathways suppresses bone nodule formation, bone mineralization, skeletal growth and bone maturation, resulting into trabecular bone loss and decrease in bone strength that leads to osteoporotic like phenotype and risk of fracture later in life.
Mercury
Mercury (
Hg) induces liver toxicity employing several processes associated with oxidative stress-mediated cell death, dysregulation of
kinases including
Gsk3 during
Wnt signaling pathways. This gluconeogenesis and adipogenesis resulted in mitochondrial dysfunction, metabolic disruption, and endocrine disruption (
83).
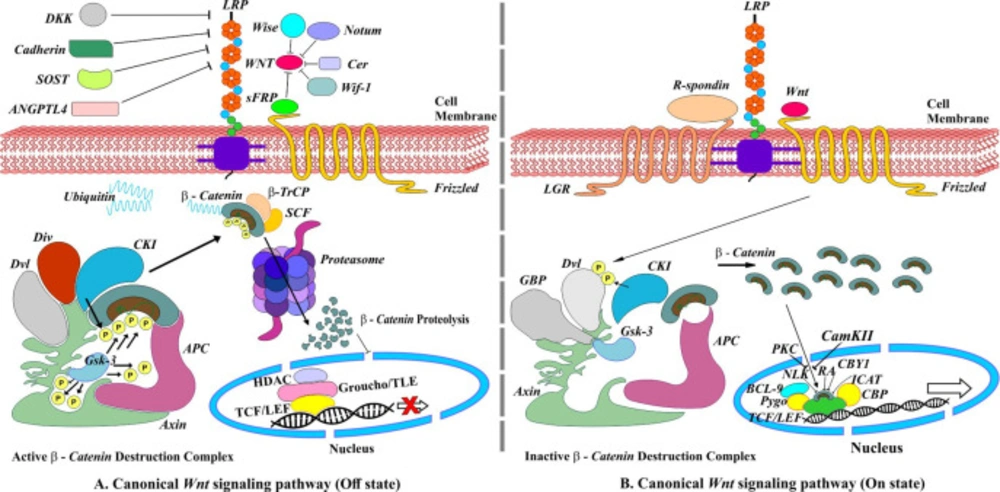
Canonical Wnt/β-Catenin signaling pathway. (A) Off state, absence of Wnt ligands leading to degradation of β-Catenin. (B) On state, the presence of Wnt ligands. R-spondins (RSPOs) is a Wnt signaling agonist that enhances Wnt signaling by binding to the members of the leucine-rich repeat-containing G protein-coupled receptor family on the cell surface
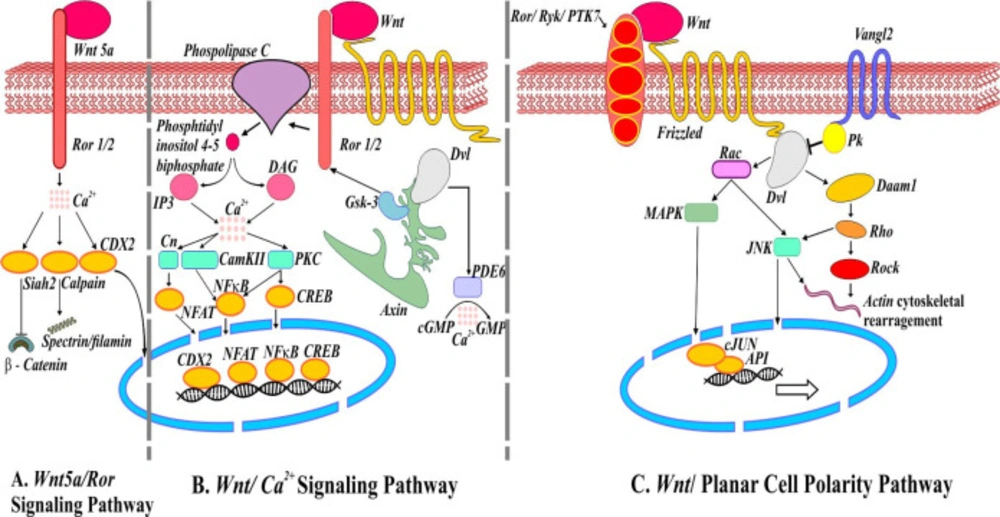
Noncanonical Wnt signaling pathways. (A) Schematic representation of mediators involved in Wnt5a/Ror signaling pathways. Activation of ubiquitin ligase Shiah2 by Wnt5a represses Wnt/β-catenin. (B) Wnt/Ca2+ signaling pathway. Wnt/Fz interaction may activate phosphodiesterase 6 (PDE6) causing Ca2+ to decrease cGMP. The release of Ca2+ induces NFAT, NFкB, and CREB translocation into the nucleus regulating the expression of genes. (C) Wnt/Planar cell polarity pathway. Van Gogh (Vangle2) forms a complex with prickle (Pk) responsible for antagonizing PCP pathway. Wnt/Fz/ Ror/Ryk/PTK7/Dvl complex also recruits Dishevelled associated activator of morphogenesis (Daam1) involve in actin cytoskeleton rearrangement
| Study (Animal/cells/tissue) | Mechanism | Outcomes | Risk | |
|---|
| Arsenic induced deregulated Wnt signaling pathways and associated risks |
| 1 | Human bronchial epithelial cells | Activates Rac1 on PKCα and Wnt5b-PKCα-mediated signaling pathway. | Activates cancer cell survival, proliferation and migration | Cancer | (27) |
| 2 | Adipose derived mesenchymal stem/stromal cells (ASCs) | Alters β-catenin levels and modulates TGFβ signaling pathway | Decreases osteogenic (Runx2, OPN and BGP) and chondrogenic (Sox9, DSPG3 and ACAN) genes expression | Induces changes in ASCs differentiation | (32) |
| 3 | Arsenic transformed cells | Activates β-catenin-VEGF pathway | Induces pro-angiogenic activity and promotes angiogenesis | Cancer | (20) |
| 4 | RIMM-18 cells | Alters Wnt/β-catenin, COX-2 and BMP signaling pathways | Decreases Wnt4, β-catenin, and BMP7 expression, increases Wt1, COX-2, MMP2 and 9 expression | Renal cancer | (18) |
| 5 | Human mesenchymal stem cells | Activates Wnt signaling pathway via upregulating Wnt3a and inhibits PPARγ, C/EBPα/β expression, and interaction between them | Reduces C/EBPs and PPARγ protein formation, inhibits adipogenesis and alters cell fate determination | Cancer | (28) |
| 6 | P19 stem cells | Repress Wnt/β-catenin signaling pathway via decreasing expression of β-catenin and other muscle and neuron-specific transcription factors | Reduces myosin heavy chain and Tuj1 expression | Inhibits myogenesis and neurogenesis | (33) |
| 7 | CRL-1807 cells | Activate ROS mediated Wnt/β-catenin signaling pathway via increased expression of β-catenin and phospho-GSK | Decreases SOD and catalase level and generation of ROS | Tumorigenesis | (22) |
| Cadmium-induced deregulated Wnt signaling pathways and associated risks |
| 8 | Mice fetus | Increases mRNA expression levels of Wnt/β-catenin target genes (Ahr, Arnt, NKx2.5, Ctnnb1 and Gsk3β) | Impairs the normal function of Ahr in regulating Wnt/β-catenin signaling during cardiogenesis, decreases total number of cardiomyocytes, swelling and apoptosis | Cardiovascular disease | (48) |
| 9 | Mice | Promote noncanonical Wnt signaling pathway and activates cdc42, increases C/EBPα while decreases Hhex expression | Impairs development of hematopoietic stem cells | Lymphopoiesistoxicity to the immune system | (45) |
| 10 | Japanese medaka embryos | Dysregulated Wnt signaling pathway via overexpression of Wnt gene, repressed bax, rad51, while inhibiting transcription of NADH-dehydrogenase nd5 gene | Increases heart rate, impairs mitochondrial respiratory chain and spinal and cardiac deformities | Teratogenicity | (49) |
| 11 | Human osteoblastic Saos-2 cells | Induces nuclear translocation of β-catenin and increased expression of Wnt/β-catenin target genes and caspase 3 activation | Induces cell proliferation and apoptosis | Bone diseases | (46) |
| 12 | Cd exposed RWPE1 cells | Dysregulated expression of ABCG2, OCT-4, and WNT-3 genes | Induces tumor growth and invasion | Oncogenic transformation | (55) |
| 13 | Chick embryo | Disrupt noncanonical Wnt/Ca2+ pathway via downregulated Wnt11, PKCα and CaMKII gene expression | Induces ventral body wall defect | Omphalocele | (54) |
| 14 | Chick embryo | Disrupt noncanonical Wnt pathway via downregulated ROCK1 and 11 gene expression | Induces ventral body wall defect | Omphalocele | (51) |
| 15 | Mice kidney | Upregulates Wnts, Fz receptors, Twist, fibronectin, collagen1 and increased expression of Wnt target genes (c-Myc, cyclin D1, Abcb1b) | Induces epithelial to mesenchymal transition that leads to renal fibrosis | Renal cancer | (58) |
| 16 | Mice kidney and liver | Dysregulates Shh and Wnt/β-catenin signaling pathway | Impairs thymocyte development | Cancer | (56) |
| 17 | BEAS-2B cells | Altered Wnt signaling pathway via upregulation of TCF4, Wnt7b and DIXDC1, UCHL1 | Initiates oncogenic transformation of lung epithelial cells | Tumorigenesis | (60) |
| Copper induced deregulated Wnt signaling pathways and associated risks |
| 18 | Zebrafish | Downregulates Wnt signaling via elevated ROS | Suppresses embryonic motility | Suppress hatching | (61) |
| 19 | Zebrafish | Downregulate Wnt signaling | Inhibits specification and formation of three swimbladder layer in a stage-specific manner | Impairs swimbladder development and inflation | (62) |
| 20 | Zebrafish | Increases canonical Wnt signaling via decreasing Wnt5 and Wnt11 transcription, altering Cmlc2, dlx3, ntl, hgg, pax2 and 6 gene expression | Smaller head, eyes and delayed epiboly | Developmental toxicity | (63) |
| Lead-induced deregulated Wnt signaling pathways and associated risks |
| 21 | Rats(Brian tissues) | Suppresses protein expression of NR2B, Arc, Wnt7a and mRNA levels of Arc/Arg3.1 and Wnt7a | Decreases spine density and dentate gyrus regions | Memory and cognitive deficit | (70) |
| 22 | MC3T3-E1 subclone 14 cells | Inactivates the Wnt/β-catenin signaling pathway by regulating Wnt3a, Dkk-1, pGSK3β and β-catenin. | Changes bone mineral composition, inhibits skeletal growth and bone maturation | Inhibits osteoblastic differentiation | (77) |
| 23 | MC3T3-E1 cells(Mice) | Depresses Wnt/β-catenin signaling due to increased sclerostin via regulating TGFβ canonical signaling pathway | Loss of trabecular bone and reduces bone strength | Osteoporosis | (74) |
| 24 | Rats | Inhibits Wnt/β-catenin pathway via reducing β-catenin, Runx2 in stromal precursor cells and increasing PPAR-γ, sclerostin protein levels | Decreases osteoblastogenesis and increases adipogenesis | Osteoporotic-like phenotype and risk of fracture | (75) |
| 25 | Mice | Inhibits β-catenin activity | Alters progenitor cell differentiation, promotes osteoclastogenesis and suppress osteoblastogenesis | Skeletal deficits | (78) |
| 26 | Rats | metal-induced | Decreases spine density and alters synaptogenesis | Impairs spine outgrowth | (79) |
| 27 | Mice | Decreases β-catenin protein along with elevated Dkk-1 and sclerostin | Inhibits endochondral ossification causing immatures cartilage in the callus | Impairs fracture healing | (81) |
| 28 | Mice | Induces TGFβ, BMP, upregulates Sox-9, type 2 collagen, aggrecan, and induces NFkappaB signaling. | Induces chondrogenesis and nodule formation | Impairs fracture healing | (82) |
| Mercury induced deregulated Wnt signaling pathways and associated risks |
| 29 | Zebrafish and human HepG2 cells | Deregulates Wnt signaling pathway, nuclear receptor and kinase activities | Triggers oxidative stress, intrinsic apoptotic pathway, gluconeogenesis, adipogenesis, mitochondrial dysfunction, endocrine disruption and metabolic disorders | Hepatotoxicity | (83) |