Many publications have highlighted the potential implications of natural products of marine macroalgae that exhibit cytotoxic activity (
10,
12). In this study, we have reported for the first time the cytotoxic activity of the CH2Cl2/MeOH (50/50, v/v) extract of the species
L. papillosa collected from the Lebanese coast. To date, numerous studies have been conducted on the cytotoxic potential of various
L. papillosa extracts on several human cancer cell lines. Especially the acetone extract of the Brazilian alga
L. papillosa which showed a moderate cytotoxic activity on several cancer cell lines, including HL-60 (leukemia), B-16 (murine melanoma), HCT-8 (human colon carcinoma), MCF-7 (human breast carcinoma), and CEM (leukemia) cancer cell lines with an IC50 of 9.9, 13.9, 15.6, 31.6, and 49.0 μg/mL, respectively (
31). In another study, no cytotoxic activity on the three breast cancer cell lines (MDA-MB-231, MCF-7 and T-47D) was detected for the ethanol extract of
L. papillosa collected from the Iranian coast of the Persian Gulf after 72 h of incubation (
32). However, the ethanol extract of
L. papillosa from the Aegean Sea in Turkey inhibited the growth of the prostate (LNCa, PC-3) and breast (MCF-7) cancer cells at 50 μg/mL, but not of the non-tumorigenic epithelial cells (MCF-10A) (
33).
Our results are consistent with those of Torres
et al. but diverge from other studies (
31-
33). Variations in the cytotoxic effect of the same algal species can be explained by the choice of extraction solvent and its ability to recover bioactive compounds, as well as by seasonal and geographical fluctuations due to changes in environmental factors such as temperature, light, and salinity (
13,
34). Regarding literature of
Laurencia, several extraction solvents were used. A mixture of CH2Cl2/MeOH seems to be relevant, as it is the most frequent used and it demonstrated ability to extract a wide range of molecules. On the whole, algae collection (geography and season), extraction and treatment conditions were optimal since our
L. papillosa extract was active compared to the previous studies.
Likewise, some metabolites such as aromatic compounds and fatty acid amide have been isolated from
L. papillosa species and their cytotoxic potential against several cancer cell lines has been previously studied. The results showed that the aromatic compounds, 4-hydroxy-benzaldehyde, and 4-methoxy-benzyl alcohol were inactive against KB cancer cells, as well as the fatty acid amide, papillamide, which did not show any cytotoxic activity against P-388 (mouse leukemia) and B-16 (mouse melanoma) cancer cells after 96 h of incubation (IC50 greater than 100 μg/mL) (
18,
20). However, most compounds isolated from
L. papillosa have not been tested for cytotoxic activity on MCF-7 human breast cancer cells. Many other reports relate to the cytotoxic activity of the compounds isolated from the genus
Laurencia against MCF-7 cells. For example, caespitol isolated from the Brazilian
L. catarinensis and teuhetenone A from the Red Sea
L. obtusa are both sesquiterpenes with high cytotoxic activity against MCF-7cells (IC50 of 9.7 and 22.8 µM respectively) (
26,
35).
Besides, UPLC-HRMS-IT-TOF (Ultra Performance Liquid Chromatography coupled with High Resolution Mass Spectrometry Ion Trap Time of Flight) analyses were performed (data not shown). Dereplication suggested the absence of already known cytotoxic metabolites in the F9 acetone fraction.
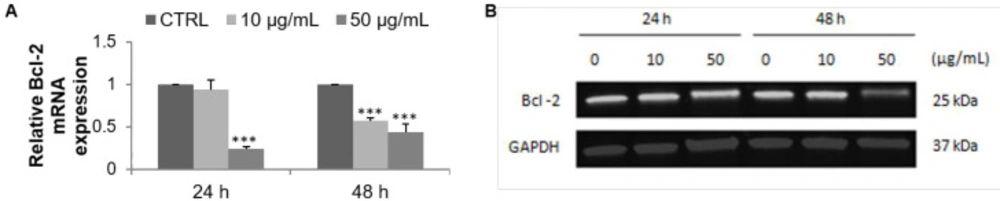
A bio-guided fractionation led to the isolation of 24 fractions. Among them, the F9 acetone fraction exhibited an important cytotoxic activity on MCF-7 cells. The cell growth inhibition can be explained by the triggering of the apoptosis process. The induction of apoptotic cell death by the F9 acetone fraction was therefore studied. The percentage of late-apoptotic cells increased while the expression of the anti-apoptotic marker Bcl-2 decreased on mRNA and protein levels. In general, apoptosis allows the normal development of the organism and the maintenance of cellular homeostasis resulting from a balance between proliferation and cell death, and any deregulation leads to cancer development and progression. Its induction causes various morphological, molecular, and biochemical changes such as chromatin condensation, DNA fragmentation, activation or inactivation of members of the Bcl-2 family, activation of the caspases, release of the cytochrome c in the cytoplasm, exposure of phosphatidylserine to the extracellular membrane, which can be estimated using various assays (
36). Our findings are in agreement with other studies showing the pro-apoptotic effect of various metabolites of algae of the genus
Laurencia, here are some examples. B16F1 melanoma cells treated with laurinterol, a halogenated sesquiterpene isolated from
Laurencia okamurai, exhibited DNA fragmentation, cell shrinkage, chromatin compaction, nuclear blebbing, and apoptotic bodies (
37). In addition, the percentage of the cells in the sub-G1 phase, the activities of different caspases, the transcriptional activity of p53, and the expression level of phospho-p53 were increased. Overall, these results indicated that laurinterol induces apoptosis via p53 activation. According to Campos
et al., elatol which is a sesquiterpene isolated from
Laurencia microcladia, was able to induce cell cycle arrest in the G1 and sub-G1 phases in B16F10 murine melanoma cells leading the cells to apoptosis (
38). The results of various biological tests demonstrated that elatol increased the number of apoptotic cells as well as the expression of the pro-apoptotic proteins p53, Bak and caspase-9, and decreased the expression of the anti-apoptotic protein Bcl-xL.
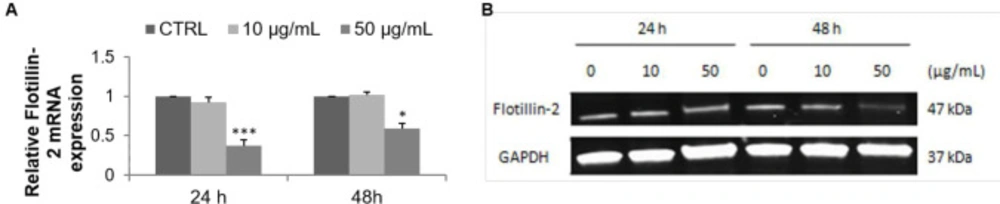
There are numerous cellular signal transduction pathways controlling the processes associated with tumor progression, such as the regulation of cell proliferation and apoptosis that occur in lipid rafts (39). These lipid rafts are defined as micro-domains within the plasma membrane enriched in cholesterol and glycosphingolipids. They serve as signaling platforms in particular for growth factor receptors such as the epidermal growth factor receptor (40). FLOT2 isolated from these micro-domains is used as a lipid raft marker protein. Its increased expression has been observed in the cancer cells relative to the normal cells, suggesting its involvement in the development and progression of various cancers including breast cancer. In addition, it has been reported that its down-regulation is associated with inhibition of cancer cell proliferation, migration, and invasion (41, 42). FLOT2 is therefore a useful biomarker and a good pharmacological target in oncology. The present study is the first to investigate the FLOT2 expression level in MCF-7 cells treated with an algal fraction. Quantitative PCR and Western blot analyses performed in this study showed a decrease in lipid raft marker expression, thus demonstrating the importance of membrane rafts as a target for cancer treatment.
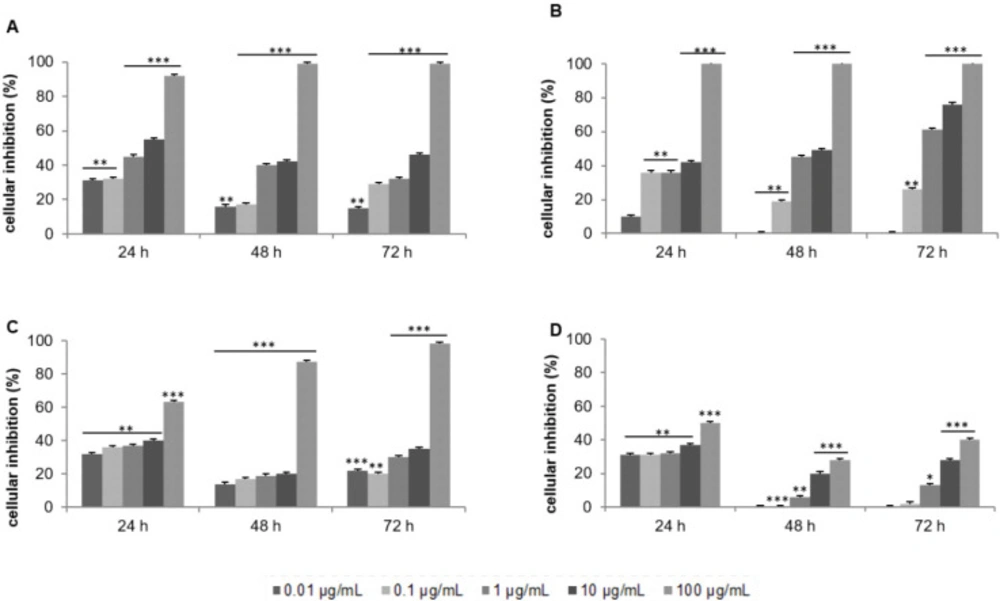
Effect of the crude extract of Laurencia papillosa and its fractions on the growth of MCF-7 cells. Cells were cultured for 24 h and treated with the indicated concentrations for 24, 48 and 72 h. Cytotoxicity was determined by MTT assay. (A) CH2Cl2/MeOH extract, (B) CH2Cl2 fractions, (C) acetone fractions, (D) MeOH fractions
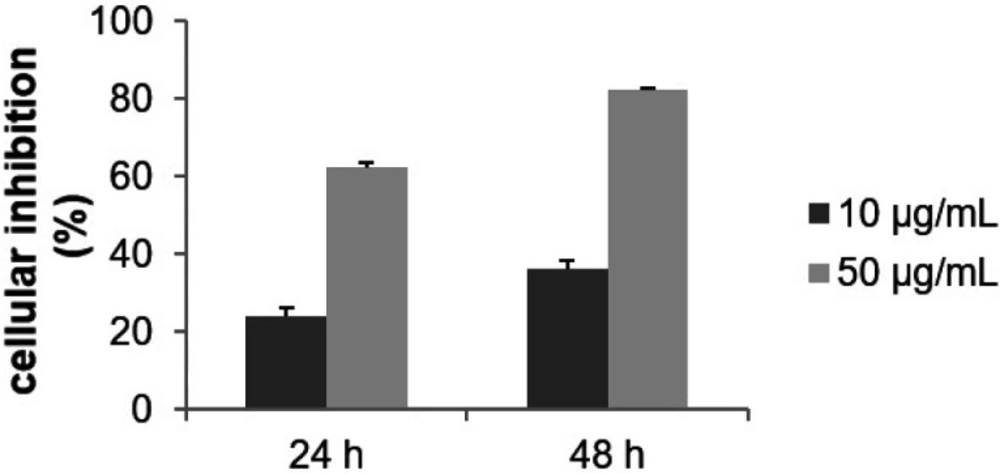
Effect of F9 acetone fraction on the growth of MCF-7 cells. Cells were treated with the indicated concentration for 24 and 48 h. Cytotoxicity was evaluated by MTT assay
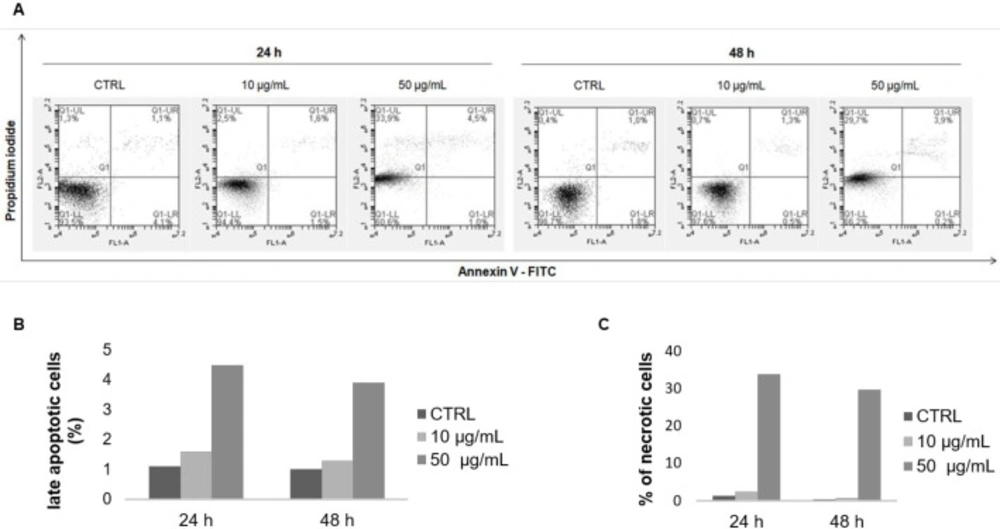
Annexin V-FITC/ PI double staining analysis of apoptosis in MCF-7 cells. Cells were treated with the F9 acetone fraction at 10 and 50 μg/mL for 24 and 48 h, and then stained with annexin V-FITC/PI and analyzed by flow cytometry. (A) Representative flow cytometry results of apoptosis: lower left quadrant, annexin V-/PI-, living cells; lower right quadrant, annexin V+/PI-, early apoptotic cells; upper right quadrant, annexin V+/PI+, late apoptotic cells; upper left quadrant, annexin V−/PI+, necrotic cells. Quantification of the percentage of (B) late apoptotic and (C) necrotic cells compared to the control group
Effect of F9 acetone fraction on apoptosis in MCF-7 cells after 24 and 48 h of treatment at 10 and 50 μg/mL. (A) RT-qPCR analysis of Bcl-2 mRNA expression, the mRNA expression was normalized to 18S and the results are expressed relative to the control set at 1. (B) Western blot analysis of Bcl-2 protein level, GAPDH is used as a control
Effect of F9 acetone fraction on Flotillin-2 expression in untreated MCF-7 cells (CTRL) and in MCF-7 cells treated at 10 and 50 μg/mL for 24 and 48 h. (A) RT-qPCR analysis, the results were normalized relative to 18S ribosomal RNA level, expressed relative to untreated cells defined as 1.00. (B) Western blot representative, GAPDH was used as a control for protein loading
| Gene | Fsorward primer sequence (5'-3') | Reverse primer sequence (5'-3') |
|---|
| 18S | GATGCGGCGGCGTTATTCC | CTCCTGGTGGTGCCCTTCC |
| Bcl-2 | GATTGTGGCCTTCTTTGAG | GTTCCACAAAGGCATCC |
| Flottilin-2 | CAAGATTGCTGACTCTAAGC | GCACAACCTCAATCTCAATC |