Natural products play an apposite role in cancer therapy today with existing numbers of anticancer agents used in the clinic being either natural or derived from natural products from various sources particularly marine organisms. Tracing for novel drugs is still important for cancer therapy, caused the fast development of anticancer drugs.
High toxicity of almost all of cancer chemotherapy drugs has triggered the request for novel anticancer drugs with little side effects and or with greater therapeutic efficiency.
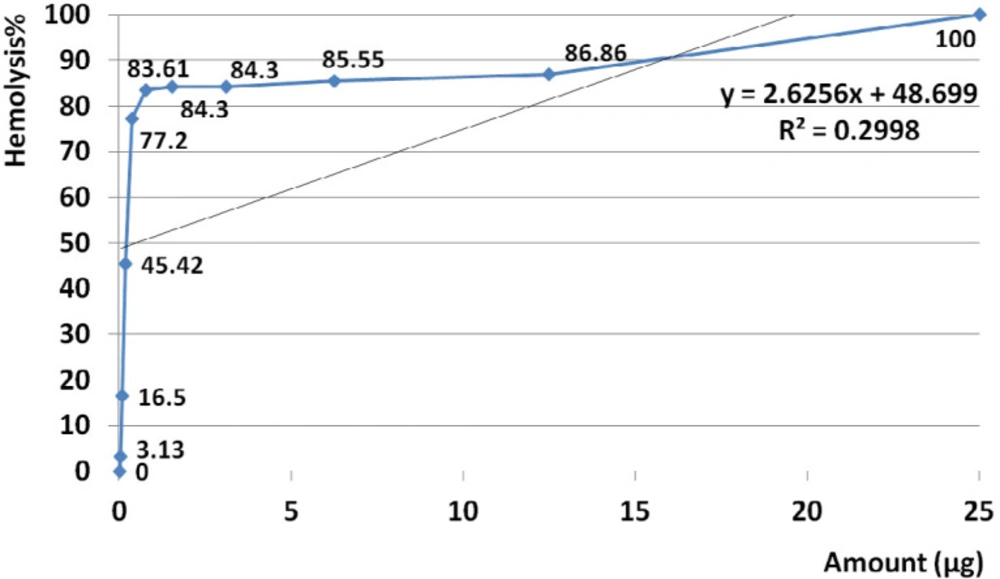
Hemolysis activity of the crude was generally not dose dependent at estimated range (R
2= 0.299) (
Figure 5). According to calculated R
2 high linearity that was seen at the range of 0.023 µg (23 ng) up to 0.39 µg (390 ng) (R
2= 0.978) indicated strange linear trend of activity. Although there is a lack of correlation between hemolytic activity and doses (R
2 = 0.3), a relative meaningful correlation about 81% is considerable to describe dose dependent trend of activity (P value 0.08). In reference to calculated R
2, high linearity was seen at examined range of doses (R
2= 0.914). This result demonstrates that hemolytic activity strangely was linear at the range of 0.78 to 25 µg.
Hemolysis activity on human erythrocytes was not seen in dose dependent manner at estimated doses (R
2=0.299) but at two distinct ranges of 0.023-0.39 µg (R
2=0.978) and 0.78-25 µg (R
2=0.914) activity was dose dependent and linearity was significant. Similarly, Subraminan
et al. in 2011 showed the hemolytic activity of
P. indicus and
P. sinensis against chicken erythrocytes and also
H. magnifica and
S. hadonii against chicken and goat erythrocytes (
41). Sudharsan
et al. in 2013 reported hemolytic activity of
S. mertensii methanolic crude venom on chicken and human erythrocytes too (
42).
Marino
et al. in 2009 showed that the crude venom from the sea anemone
Aiptasia mutabilis had dose-response hemolytic activity against human erythrocytes (
43) but our results disagree with that.
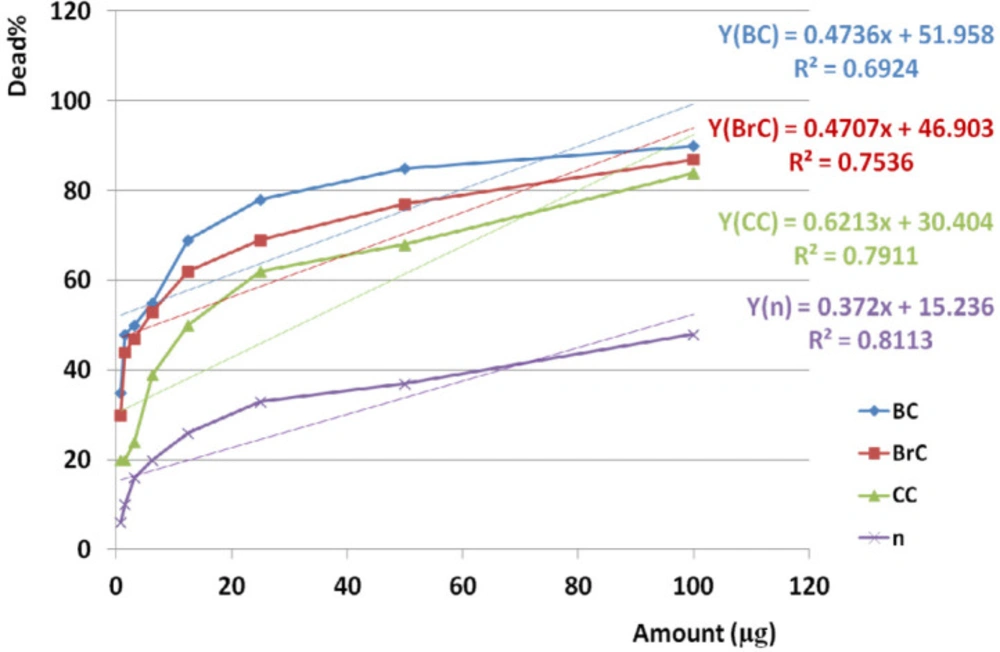
Amazing Similar activity of crude venom on BC, BrC, and CC cell lines based on variance analysis (p value <0.001), not only may give us a chance to finding an anticancer fraction but may boost our chance to apply lower doses of anticancer agent and avoiding toxicity in normal cell line. The results obtained from Cross comparison of anticancer activities of crude venom between BC and BrC, BC and CC, BrC and CC cell lines showed that each examined cancer cell line has different sensitivity when exposed to the same crude venom. Crude venom induced more lethality on BC cells than the other cancer cell lines. Significant difference in toxicity of crude venom against examined cancer cell line compared to normal cell line (p value <0.001) is an interesting phenomenon pointed out to existence of a non-toxic anticancer fraction in crude venom. According to significant cross-correlations between anticancer activity of crude venom on BC and BrC, BC and CC, BrC and on CC cell lines, similar mechanism of toxicity would be contemplated. In feference to significant correlation between anticancer activity of crude venom on BC, BrC, and CC with NHDF cell lines by increasing examined doses, this similarity would suggest a common dose dependent trend of toxicity on all three cancer cell lines and normal cells as well.
Cline
et al. in 1995 pointed out cytotoxic effect of crude extract from the sea anemone
Urticina piscivora and showed that the growth of 50% of oral human epidermoid carcinoma cells (KB), mouse lymphocyte leukemia cells (L1210), and human embryonic lung diploid cells (HEL299) were inhibited at 6.54, 10.07, and 2.34 mg/mL, respectively (
44). Our study also demonstrated anticancer activity of
S. haddoni crude extract.
The crude venom extracted from
H. malu,
C. adhaesivum and
E. quadricolor had a significant inhibitory effect on skin cancer, A431 cells. Furthermore,
H. malu and
C. adhaesivum had a significant inhibitory effect on breast cancer, T47D cell line at 40 μg/mL (
25). This result was in accordance with toxicity of
S. haddoni crude venom on breast cancer cell line, MDA-MB231 approximately.
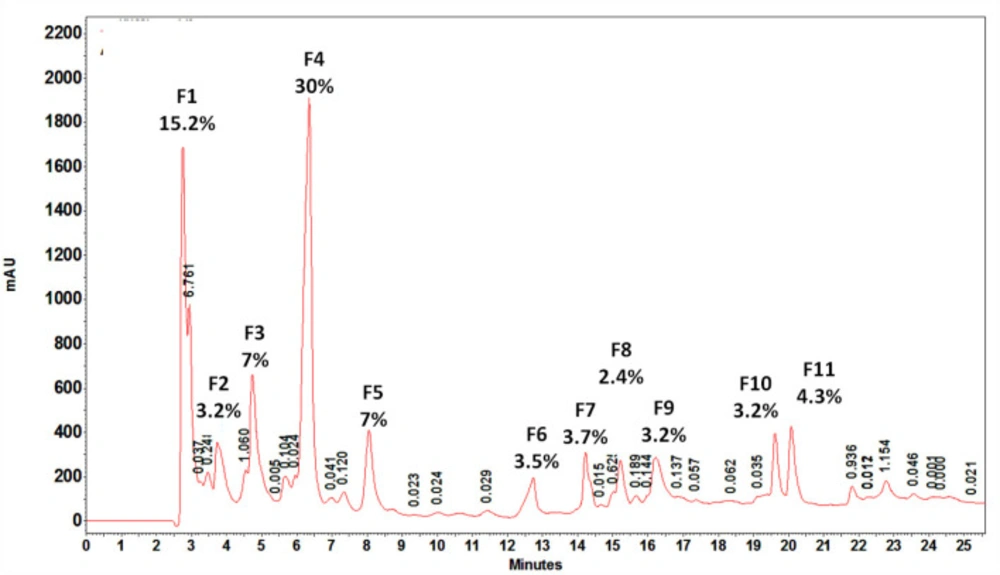
According to observed retention time, almost all of the fractions were relatively hydrophobe and eluted from 1.357 to 17.8% acetonitrile. The first three fractions were eluted at 2.758, 3.732, and 4.739 min referring to hydrophilic proteins or peptides. The largest peak was seen at 6.357 min representing a very weak hydrophobic fraction eluted at 1.357% acetonitrile.
According to significant correlation between hemolysis activity of all hemolytic fractions and examined doses, activity was indicated as dose dependent and a linear trend of activity was showed. Similar trend of hemolysis would be indicated to the same mechanism induced by the same hemolytic agent.
According to analysis of variance for all hemolytic fractions, similar activity at all doses of each fraction was showed (p value < 0.05). Comparing the mean activity of all fractions together by paired sample t-test showed significant difference between them (p value <0.05) except for the following pairs including F1-F2, F1-F3, F2-F4, F2-F5, and F5-F6 (p value >0.05).
Analysis of correlation between hemolysis activity and examined doses by linear regression test indicated that all fractions had dose dependent manner. Cross comparison of trend of hemolysis in all hemolytic fractions was performed by linear regression test and the results showed similar trend of activity in all fractions (R2 = 0.74-0.999).
Ravindran in 2010 showed hemolytic activity in all fractions isolated from three sea anemone including
Heteractis magnifica,
S. haddoni, and
Paracodylactis sinensis collected from Indian costal region. Interestingly, some of our HPLC isolated fraction had no hemolytic activity (
45).
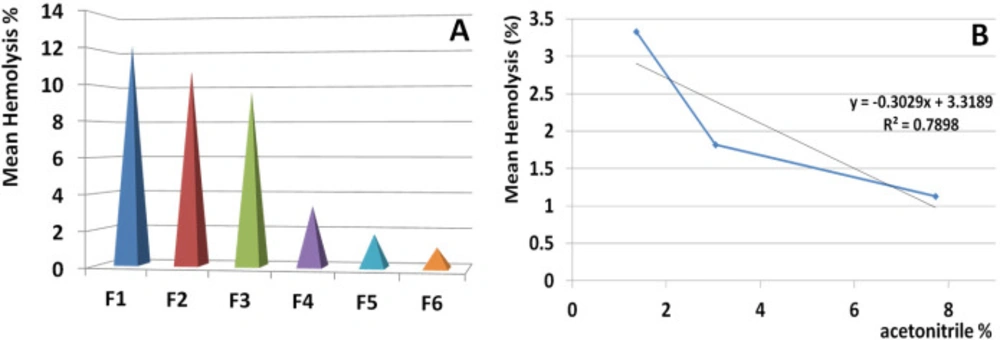
During data interpretation regarding hemolytic activity of the isolated fractions, a hypothesis was triggered concerning the correlation of hemolytic activity and hydrophobicity. Following further analysis by linear regression test, a significant correlation was showed between activity of hemolytic fraction and the percent of acetonitrile in which the fractions were eluted.
This issue indicated that hydrophobic fractions have lower toxicity against human RBCs. Interestingly the first three hydrophilic fractions had greater toxicity than the subsequent weak hydrophobic fractions demonstrating that hydrophobicity of S. haddoni fractions is in accordance with no hemolytic activity.
Analysis of variance for anticancer activity of isolated fractions on cancer cells showed that activity of each dose of each fraction was generally similar together (p value of all fractions: < 0.01).
Cross comparison of the mean of anticancer activity of each fraction with the other fractions by Paired sample
t-test showed significant differences in almost all isolated fractions against BC, BrC, and CC (
p value < 0.05) in exception of some fractions. In reference to the difference of hydrophobicity in isolated fractions (
Figure 6) and difference in their anticancer activity, it could be suggested that each isolated fraction has a distinct entity or mechanism.
Anticancer activity of examined doses of all fractions against breast, brain, and colon cancer cells were correlated significantly together by linear regression analysis (R2= 0.528 - 0.95).
Significant difference between toxicity of F9, F10, F11 fractions on normal cell line pointed out to distinct entity and to different mechanism of toxicity.
According to significant difference between toxicity of F9, F10, and F11 on normal and cancer cells, F10 was selected as our candidate fraction since it had the lowest toxicity on normal cells.
Comparison of trend of toxicity for F9, F10, and F11 fractions showed a similar behavior and this issue suggested that these three fractions have possible similar mechanism of toxicity.
Since the mean toxicity of F10 on the cancerous cells is remarkably upper than the normal cells, dose dependency of F10 provided us with a good chance to access a non-toxic dose on normal cell. Furthermore, it means that lower dose of F10 at nanomolar scale avoiding immune system to react against the molecule as immunogen.
Cline
et al. in 1995 demonstrated anticancer effect of a protein designated UpI isolated from the sea anemone
Urticina piscivora and showed that the growth of 50% of oral human epidermoid carcinoma cells (KB), mouse lymphocyte leukemia cells (L1210), human embryonic lung diploid cells (HEL299) were inhibited at 40.32, 29.99, and 29.74 mg/mL respectively (
44). UpI was found to be hemolytic on human RBC but hadonin showed no hemolytic activity in our study.
In reference to Jiang
et al. study in 2003, a recombinant protein, Src I, from the sea anemone
Sagartia rosea exhibited 50% cytotoxic activity on human cell lines, including astrocytoma (U251) (3.5 μg/mL), Non-Small Cell Lung Cancer Carcinoma (NSCLC) (2.8 μg/mL), liver carcinoma (BEL-7402) (3.6μg/mL), stomach adenocarcinoma (BGC-823) (7.4 μg/mL), NIH Swiss mouse embryo (NIH/3T3g) (3.4 μg/mL) (
34). Src I as an anticancer agent had been showed hemolytic activity and in fact, was found to be strongly toxic (HD50 at 0.43 μg/mL) but hadonin showed no hemolytic activity in our study.
Avila
et al. in 1988 isolated a toxin from the sea anemone
Stoichactis helianthus that exhibited toxicity on HL-60 human myelocytic leukemia cells (
46) but also was toxic against human erythrocytes and human peripheral mononuclear cells too while hadonin induced no toxicity on human RBCs and human normal fibroblast cells as well.
Fedorov
et al. in 2010 isolated an anticancer protein, RTX-A, from the sea anemone
Heteractis crispa. RTX-A reduced cell viability of JB6 P+ Cl41 cells, Hela, THP-1, MDA-MB-231 and snu-c4 human tumor cell lines (
33).
EqTX-I cytolysis toxin from the sea anemone
Actinia equina also induces a decrease in the viability of V-79-379 A cells (diploid lung fibroblast from Chinese Hamster) in a concentration-dependent manner (
47) while hadonin showed exceptional non-toxic activity on human normal fibroblast cell line.
EqTx-II from
Actinia equina was studied for cytotoxicity against human glioblastoma U87 and A172 cell lines. After 24 h of treatment, 10 mg/mL EqTx-II showed remarkably cytotoxic and reduced the viability of U87 and A172 cells to 60 and 48%, respectively, but was toxic on normal cells at 10 mg/mL and decreased the viability to 80% (
27) while hadonin showed anticancer activity on the examined cancer cell lines but non-toxic activity on human normal fibroblast cell line at a dose of 20 ng.
According to obtained results, anticancer activity of hadonin is of high pharmaceutical value to follow its therapeutic potency in animal models.
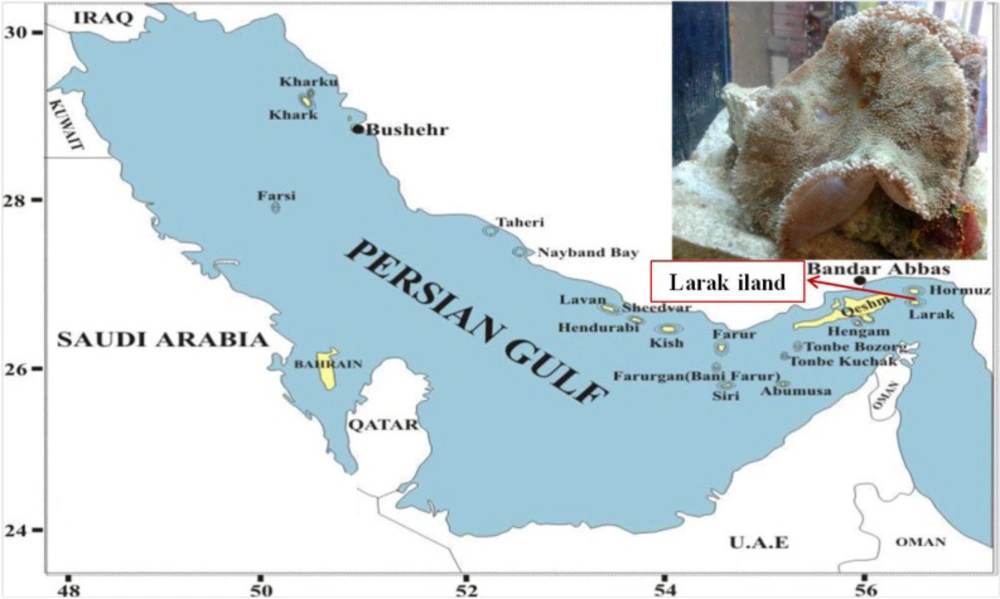
Specimen collection area. Larak Island, the Persian Gulf (26°51'12" N 56°21'20" E).
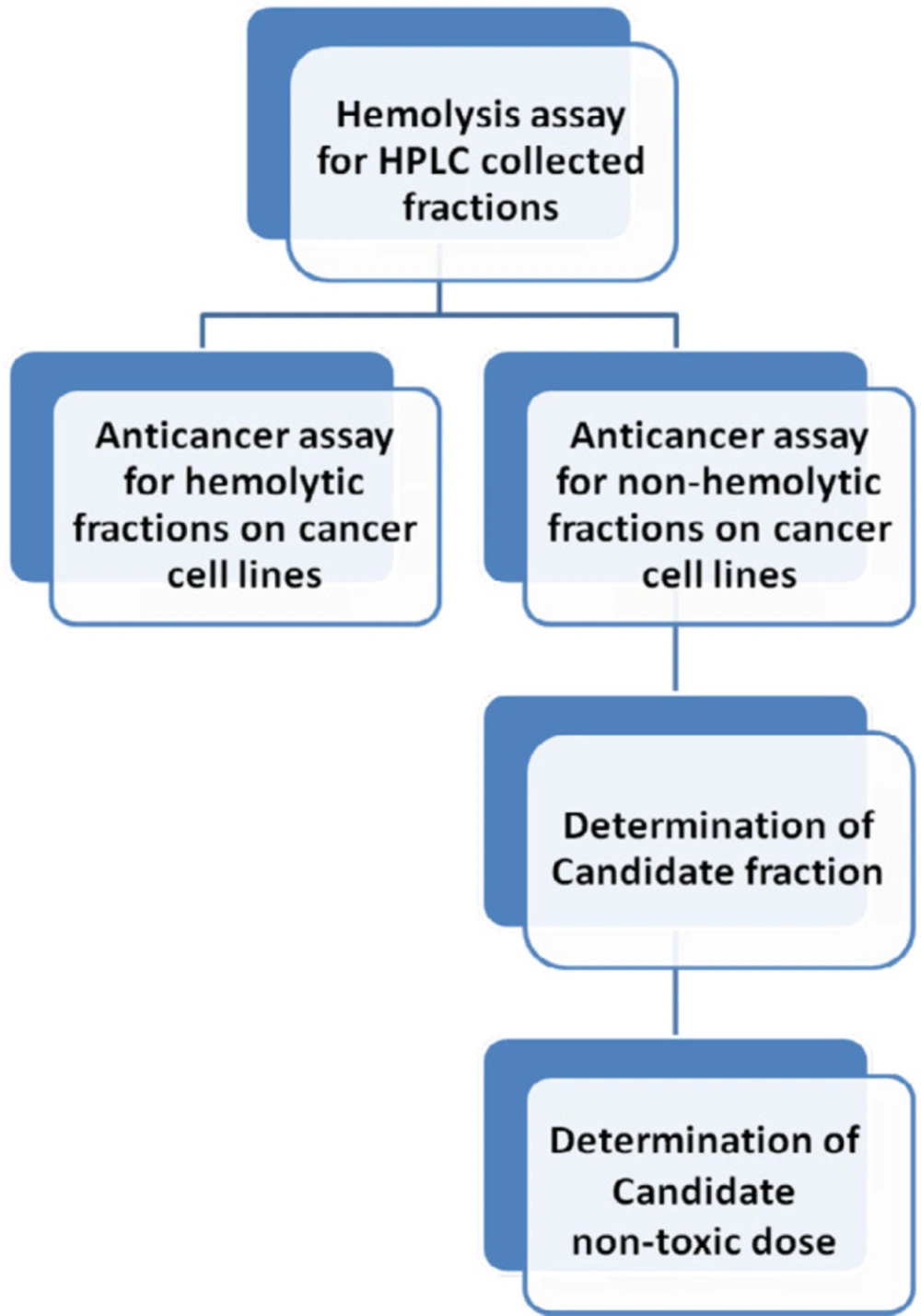
Demonstration of classified goals to reaching a non-hemolytic non-toxic anticancer fraction
Macroscopic and microscopic image of the tentacles of Stichodactyla haddoni. A. Macroscopic image. B. Magnified lateral view of tentacles. C. Microscopic image of a tentacle containing capsule and barb (C: 40X).
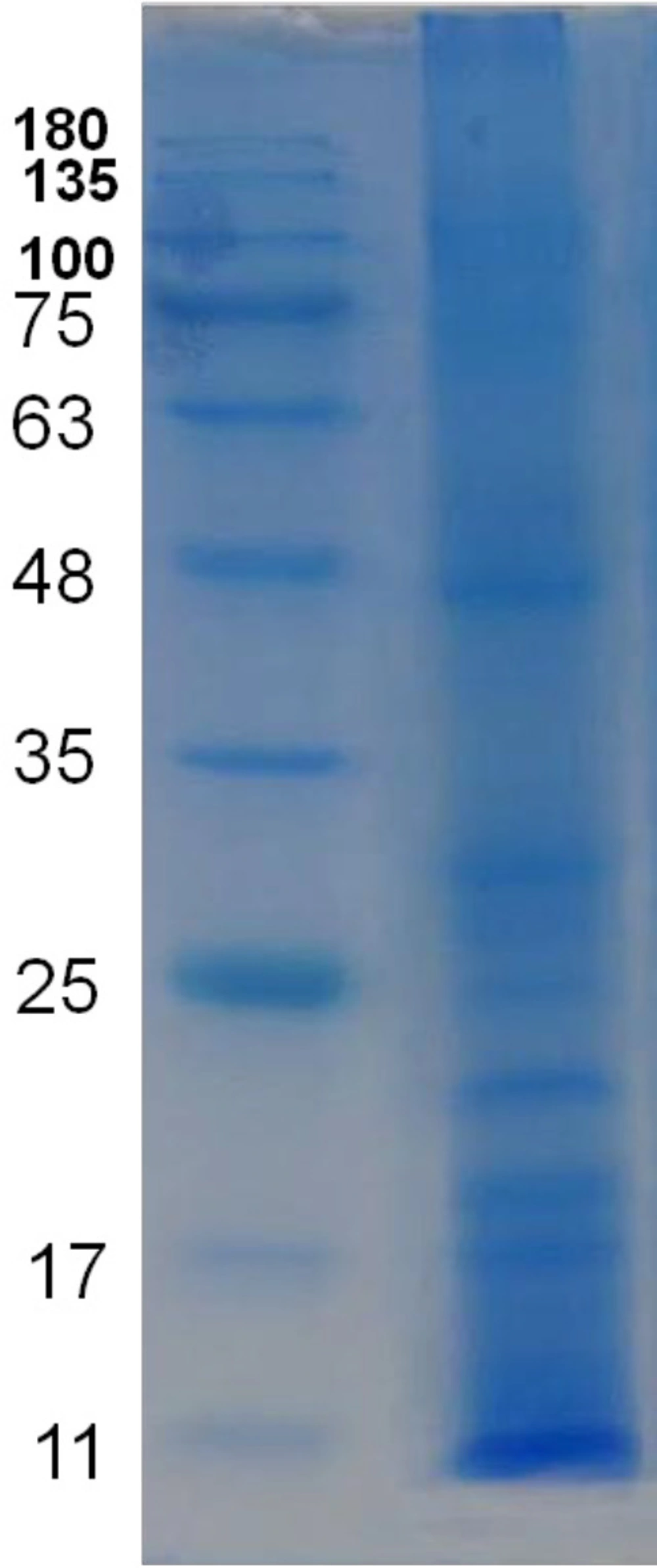
Protein profile of water extract of the venom of S. haddoni. SDS-PAGE results showed about 12 separate bands and molecular weight of observed proteins ranged approximately from 8 to 250 kDa
Hemolytic activity of water extracts S. haddoni crude venom. The amount of 25 µg crude venom produced 100% hemolysis and HD50 identified at 0.5 µg. Based on regression analysis, no general linearity was seen at examined amounts of venom ranged from 0.023 to 25 µg. This indicates that hemolysis activity was generally not dose dependent at estimated range (R2= 0.299).
Anticancer activities of the crude extract against cancer and normal cell lines during 24 h. EC50 of crude venom against cancer and normal cell lines was observed at 4.13, 6.58, 31.54, and 93.45 µg respectively. Linear regression analysis showed significant correlation between anticancer activity of crude venom on BC, BrC, and CC and n cell lines and examined doses (R2BC = 0.692, R2BrC = 0.754, R2CC = 0.791, R2n = 0.811). “BC”, “Brc”, “CC”, and “n” are abbreviations of Breast Cancer, Brain Cancer, Colon Cancer, and Normal cell line
HPLC profile of water extract of tentacles removed from S. haddoni. 11 major and minor peaks were seen and area percent greater than 3% collected to further investigations. F4 had the greatest are percent
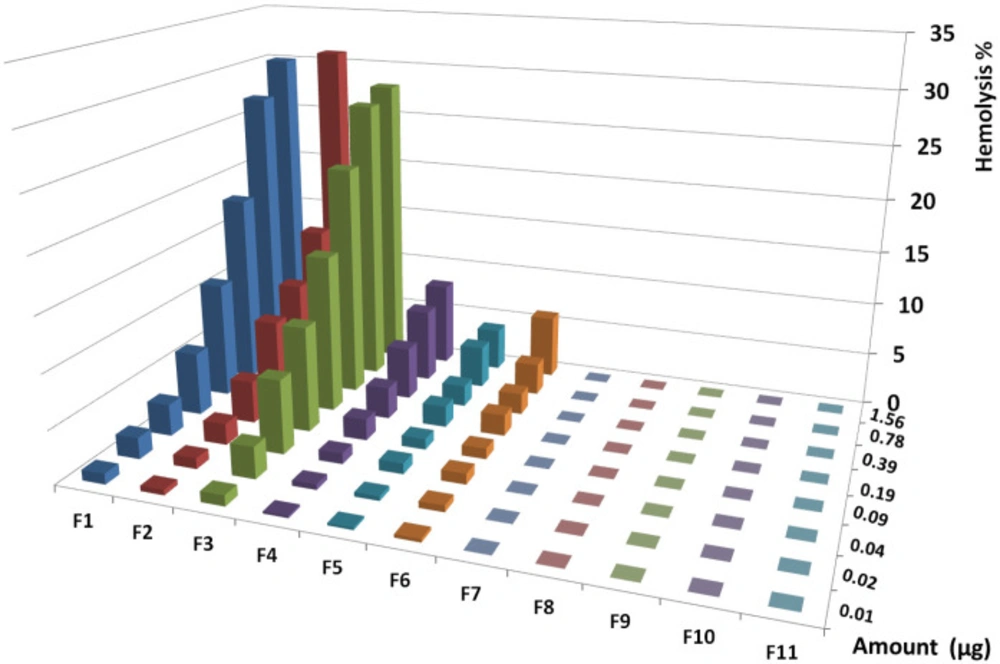
houreshemolytic activity of collected HPLC fractions on human RBCs
Correlation of hemolytic activity and hydrophobicity of isolated fractions. Linear regression test was performed to control the correlation between activity of hemolytic fraction and the percent of acetonitrile in which the fraction eluted. The result indicated a meaningful correlation (R2 = 0.789). This issue indicated that hydrophobic proteins have the lowest toxicity for human RBCs and also the hydrophobicity is in accordance with no hemolytic activity
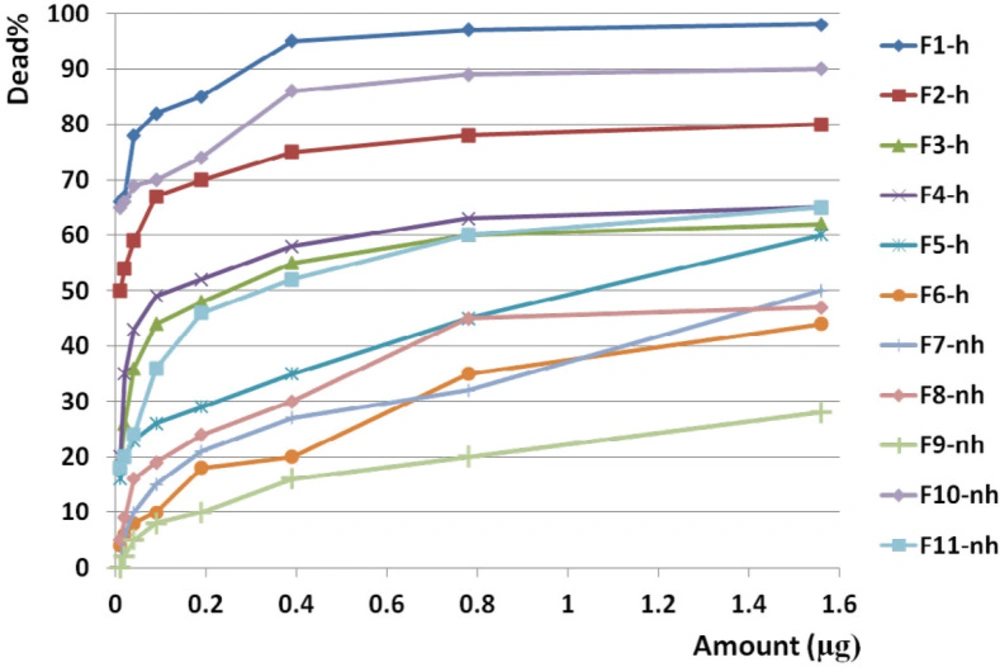
Anticancer activity of all fractions from F1 to F11 on breast cancer cell line. Anticancer activity of examined doses of all fractions against breast cancer cells were correlated significantly together by linear regression analysis (R2= 0.529 - 0.95). Cross comparison of the mean of anticancer activity of each fraction with the other fractions by Paired sample t-test showed significant differences in almost all isolated fractions against BC cells (p value < 0.05).
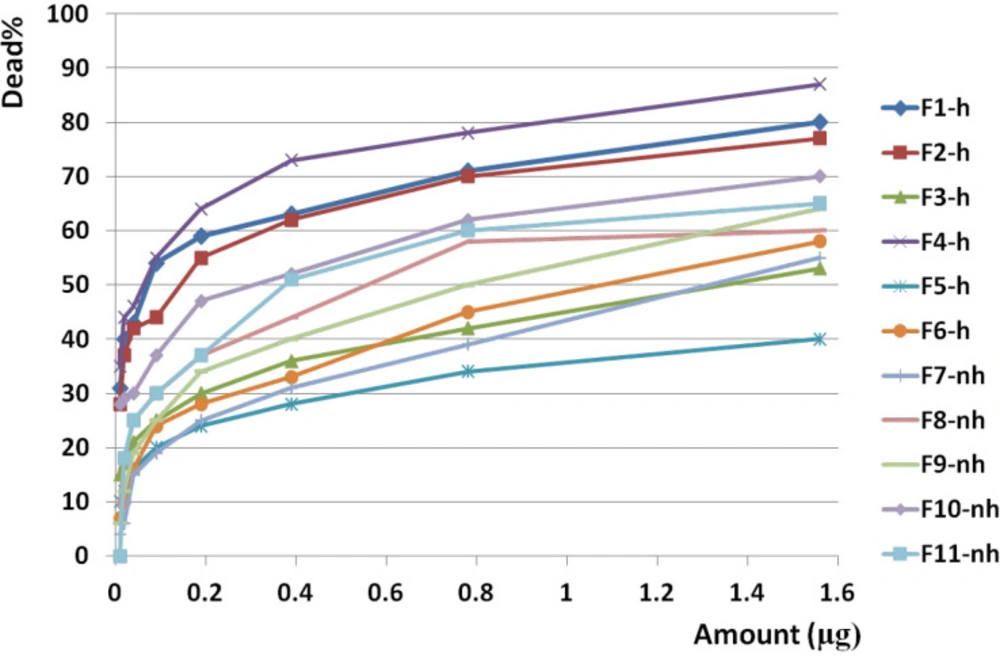
Anticancer activity of all fractions from F1 to F11 on brain cancer cell line. Anticancer activity of examined doses of all fractions against brain cancer cells were correlated significantly together by linear regression analysis (R2= 0.679 - 0.868). Cross comparison of the mean of anticancer activity of each fraction with the other fractions by Paired sample t test showed significant differences in almost all isolated fractions against BrC cells (p value < 0.05).
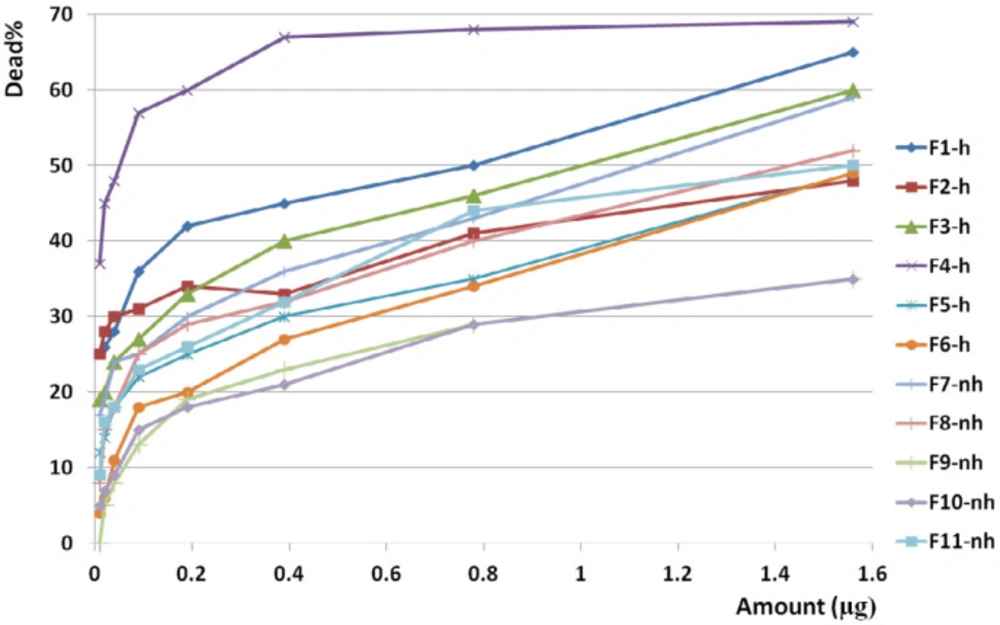
Anticancer activity of all fractions from F1 to F11 on colon cancer cell line. Anticancer activity of examined doses of all fractions against colon cancer cells were correlated significantly together by linear regression analysis (R2= 0.528 - 0.945). Cross comparison of the mean of anticancer activity of each fraction with the other fractions by Paired sample t-test showed significant differences in almost all isolated fractions against CC cells (p value < 0.05)
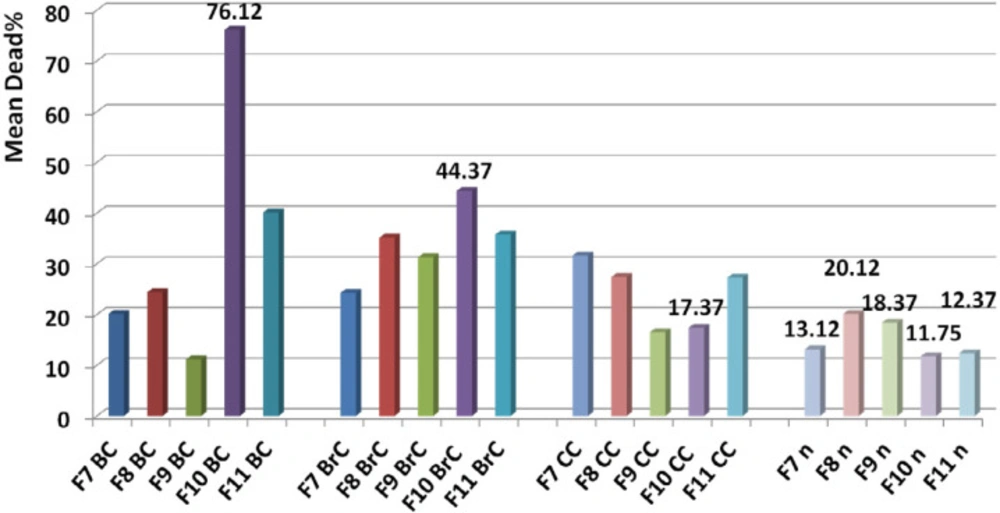
Determination of candidate fraction. Comparison of toxicity of F9, F10, and F11 on normal cell with the other cancer cells showed a significant difference based on paired sample student t-test (P value < 0.05) while F7 and F8 had similar toxicity with F10 and F11, and F9 respectively (P value > 0.05). Comparing the mean toxicity of the fractions between together showed that F10 had the least toxicity and selected as candidate fraction
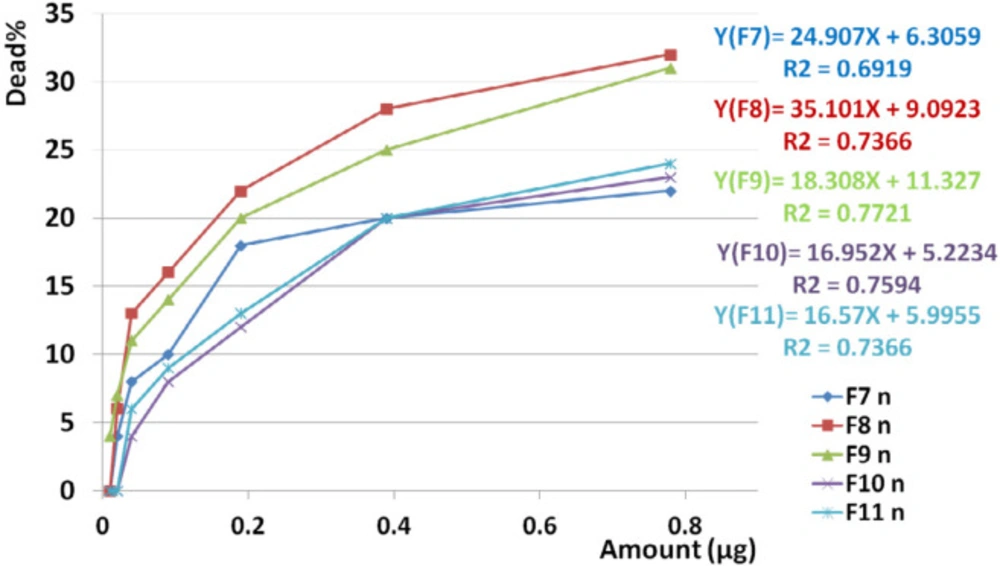
Toxicity of non-hemolytic fractions on normal cell line. Linear regression analysis showed significant correlation between toxicity of F7, F8, F9, F10, and F11 on normal cell lines with increasing doses of the fraction (R2F7 = 0.7, R2F8 = 0.73, R2F9 = 0.772, R2F10 = 0.759, R2F11 = 0.736).
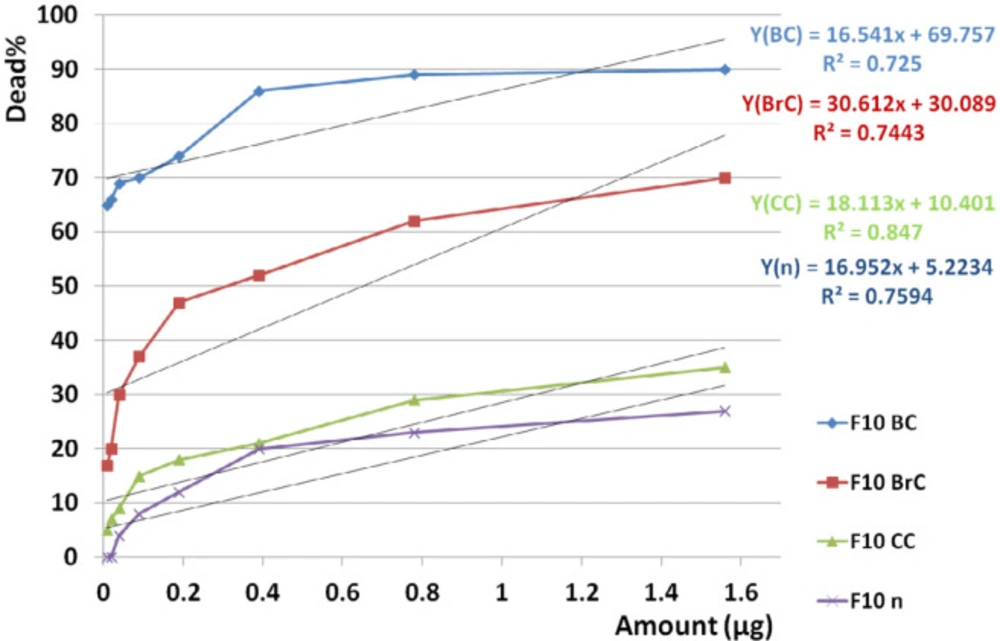
Dose dependency of the candidate fraction, F10, on cancer and normal cell lines. According to linear regression analyses, toxicity of F10 on all three cancer cell lines was dose dependent (R2BC = 0.725, R2BrC = 0.744, R2CC = 0.847) as well as on normal cell line (R2n = 0.759). It means that with decreasing the doses, there is a good chance to access a non-toxic dose on normal cell since the mean toxicity of F10 on cancerous cell is remarkably upper than normal cells
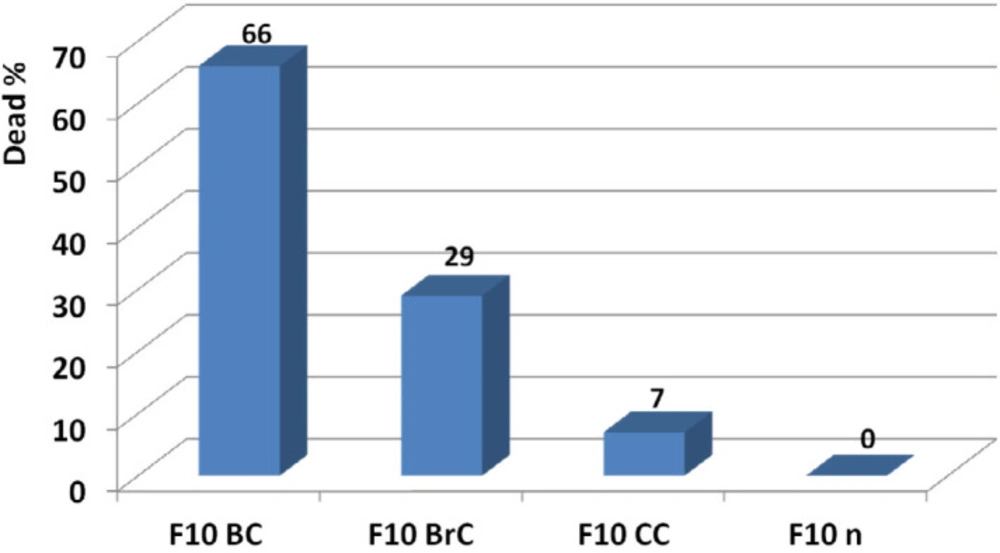
Determination of effective candidate non-toxic dose of F10 and comparison of its activity on breast, brain, and colon cancer cell lines. According to results, F10 showed the best anticancer activity on breast cancer cell line at 20 ng. At this amount no toxicity was observed on normal fibroblast human cell line
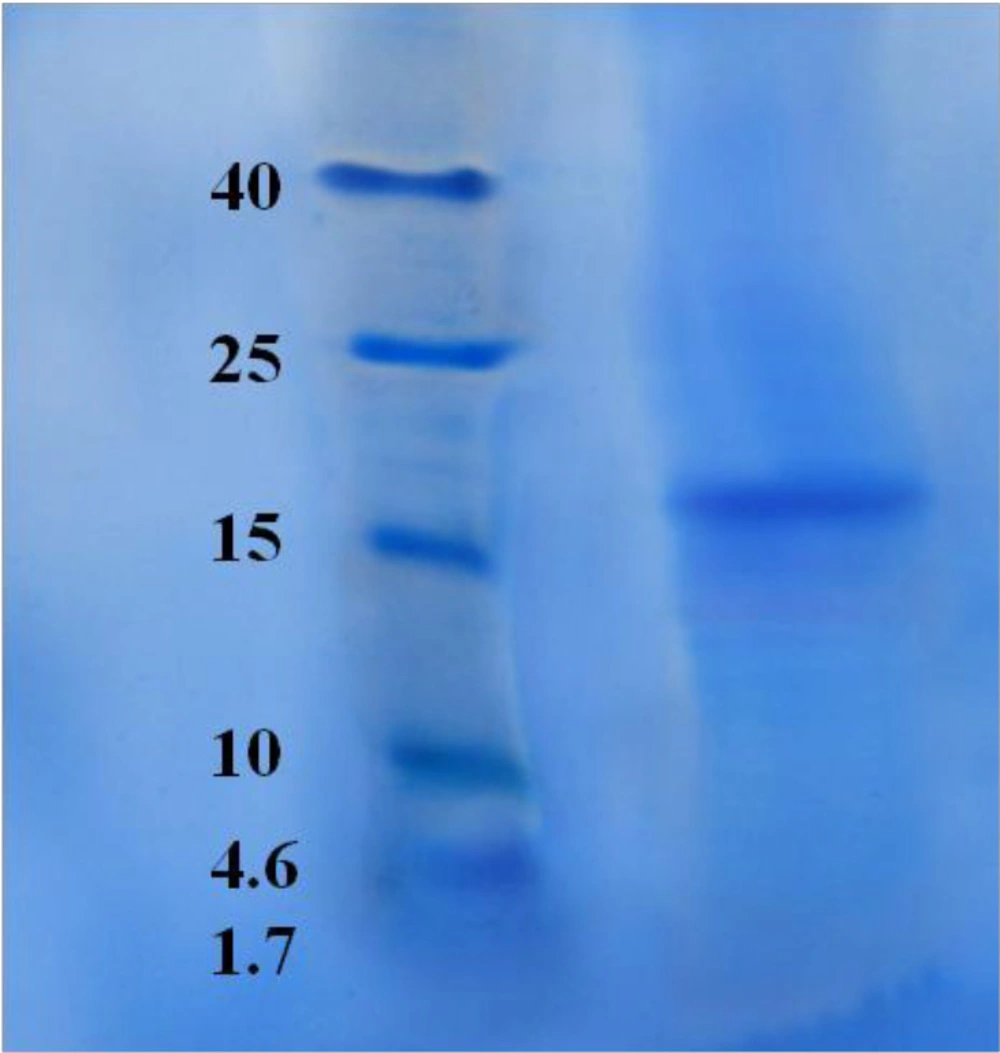
SDS-PAGE for HPLC purified candidate fraction, F10. SDS-PAGE showed one pure band corresponding to molecular weight approximately at 17.5 kDa