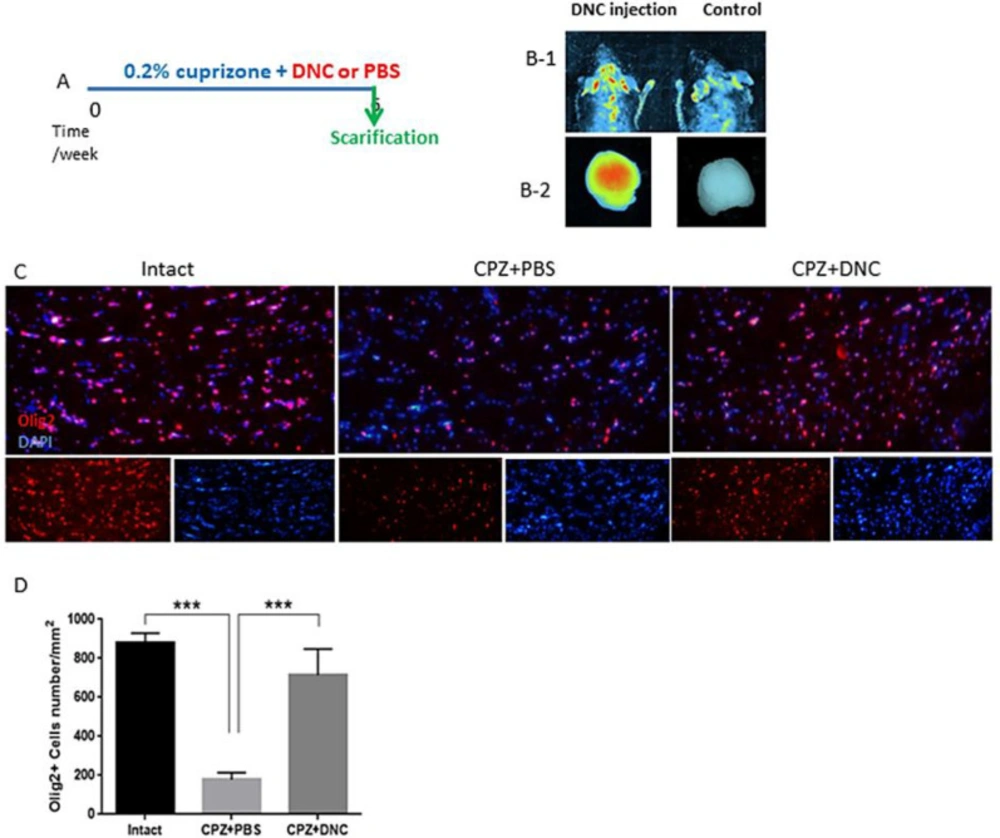
DNC ability to cross the intact BBB
The presence of the BBB, a highly selective semipermeable border between the capillaries and the brain is a major obstacle that dramatically hinders the drug delivery. Available reports are controversial for integrity of BBB in CPZ fed mice. Here, to ensure that DNC is able to pass intact BBB we used an
in-vivo imaging system. Our observation indicated that DNC is detectable in brain region of mice with intact BBB within 1-2 h after being injected intraperitoneally (12.5 mg/kg). We also could detect DNC fluorescence emission in harvested brain, under UV-light excitation beam. However, with the same exposure time and gain settings, no significant fluorescent signal was indicated in mouse that did not receive DNC injection (
Figure1, B 1-2).
The Effect of DNC on CPZ-induced changes in the OLLC composition of CC
CPZ demyelination is induced by oligodendrocytes toxic death, rather than an attack on the myelin directly. Reactivation of astrocytes, microglia accumulation, mitochondrial injury, and release of ROS have been suggested as a major driving force of OLs apoptosis. Thus, approaches that modulate these mechanisms potentially provide a new range of therapeutic strategies. Traditionally, OLs death and demyelination are viewed as targets of an uncontrolled autoimmune inflammatory response. But, a number of clinical and experimental experiments have reported that OLs injury/death seems to occur prior to disease process and initiate the formation of demyelinating lesions (
1,
31,
32). However, it is suggested that protection of OLs against injury results in protection against MS progression and symptoms (
33,
34). On the basis of curcumin-mediated cytoprotection against oxidative stress and inflammation (
35,
36), here we investigated that protective effect of DNC on OLLCs in context of CPZ toxicity. Thus, the numbers of OLLC was investigated by IHF against Olig2, as specific marker of these cells. In CPZ + PBS group, CC contained reduced numbers of olig2
+ cells compared to the control; however, the number of Olig2
+ cells was significantly higher in CPZ+DNC treated mice compared to PBS treated group (
Figures1 C-D). As CPZ is known to induce specific death of OLLC, this result suggested that DNC exhibited protective effect on these cells against CPZ. Another explanation might be the inductive effect of DNC on migration of CNS resident oligodendrocyte progenitor cells (OPCs) toward demyelinated lesions.
Available studies have reported the neuroprotective effect of curcumin or its nano-formulations in different CNS disorders. This effect is well-established to be at least in part via anti-inflammatory and anti-oxidative stress property of curcumin. I one available study, curcumin is reported to protect pre-oligodendrocytes from infection-derived apoptosis. This protection is suggested to be associated with suppression of nitric oxide synthase (iNOS) and NADPH oxidase (NOX) (
37). However, to best of our knowledge no available study has reported protective effect of curcumin or its nano-formulations in demyelinating lesions.
The Effect of DNC on astrocytes and microglia composition of CC
Astrocytes reactivation and microglia accumulation are well-established features of demyelinated lesions in MS. Both of these events lead to production of large amount of ROS, leading to oxidative injury followed by mitochondrial dysfunction. Thus, they play significant role in OLs injury and their dysfunction.
As CPZ-induced demyelination is accompanied by activation and accumulation of astrocytes and microglia, mimicking a central feature of MS, we investigated the effect of DNC on number of astrocytes and microglialcells within CC by IHF against GFAP
+ and Iba1
+ as markers of these cells, respectively. As shown in
Figures 2 A-B, the CPZ+PBS group showed significantly increased number of GFAP
+ cells compared to the control group; however, DNC treatment was associated with significantly lower number of GFAP
+ cell in CC, when being compared with PBS treated mice. Astrogliosis formation is one of the consequences of myelin damage; thus, this finding suggested inhibitory effect of DNC on glial scar formation. Accumulation of reactive astrocytes at the site of CNS injury is a seminal feature of damaged or diseased CNS. As, this impairs OLLS survival and remyelination capacity; we suggest that DNC may exhibit its protective effect on OLLCs by reduction of astrocytes’ activation.
As mentioned, CPZ model coincide with Iba-1 + microglia migration toward CC. Accumulated microglia release inflammatory and cytotoxic mediators, including ROS and iNOS. This contributes to the formation of inflammatory microenvironment, cell death and neurological dysfunction. OLs are the main target cells in such cytotoxic condition. It is reported that the number of Iba-1+ microglia is inversely proportional to OLs, suggesting that the accumulation of inflammatory microglia is closely related to myelin loss. As we have previously showed anti-oxidative stress and anti-inflammatory properties of DNC, here we investigated the direct effect of this compound on accumulation microglia, as main producers of inflammatory mediators. Our results showed that even though the number of microglial cells was significantly higher in CPZ+PBS treated groups, CPZ feeding accompanied with DNC treatment lead to significantly lower number of these cells in CC (
Figure 2C-D).
Pathological process including inflammatory response and glial scar formation are major causes of progression of CNS- injuries and inhibition of repair. Curcumin has been showed to curcumin promotes spinal cord repair through inhibiting glial scar formation and inflammation (
38). Under the influence of many relevant factors, microglia cells and astrocytes are activated in features of MS lesions. This will lead formation of dense glial scar and production of excessive amount of inflammatory mediators and extracellular matrix components, such as chondroitin sulfate proteoglycan. This process leads to death of myelination cells and inhibition of remyelination. Our previous study, along with the other reports has indicated neurotrophic property of curcumin and its nano-formulation, in CNS damage context (
9). As a neurotrophic environment contributes to the survival and regeneration of OLLCs in CPZ-induced demyelination, we suggest that DNC promoted OLLCs survival/proliferation by providing a support to inhibit their apoptosis by CPZ.
The effect of DNC on myelination index of CC following acute CPZ intake
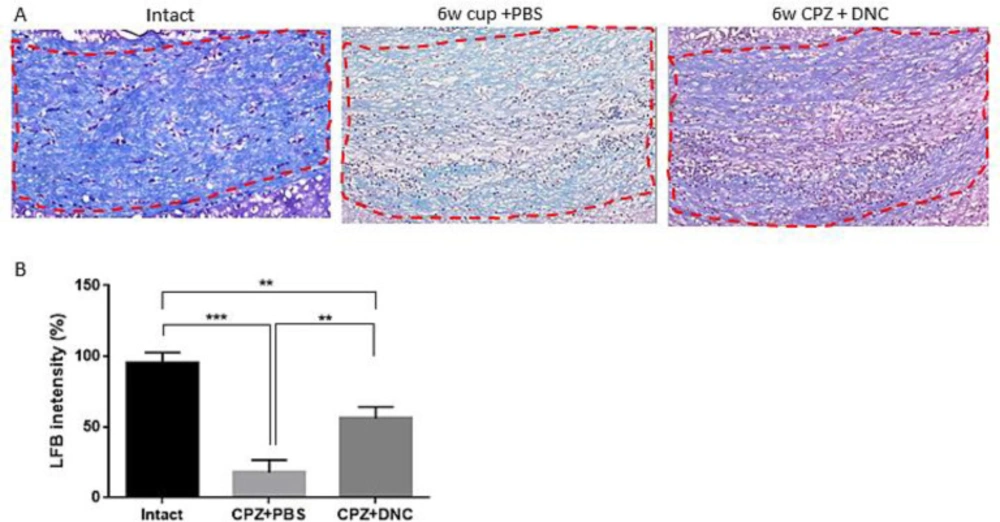
Considering protective effect of DNC on OLLC, along with its repressive effect on astrocyte and microglia activation, hypothetically we assumed that DNC protects against toxic myelin damage by CPZ. In this regard, we evaluated the protective effect of DNC on myelin content of CC. It is well-established that after 5-6 weeks CPZ intake, the CC region losses its myelin content and an acute demyelination occurs. Hereby using LFB staining, we observed a normal myelin structure in the intact group (
Figure 3A). However, the animals treated with CPZ and PBS (CPZ+PBS) exhibited a significant decrease in LFB staining, characterized by light and a disorganized myelin sheet in CC. Comparing the semi-quantitative results of LFB intensity showed that in the animals treated with CPZ and DNC (CPZ + DNC group), the degree of LFB staining was significantly higher than that in the CPZ + PBS group (
Figure 3A-B).
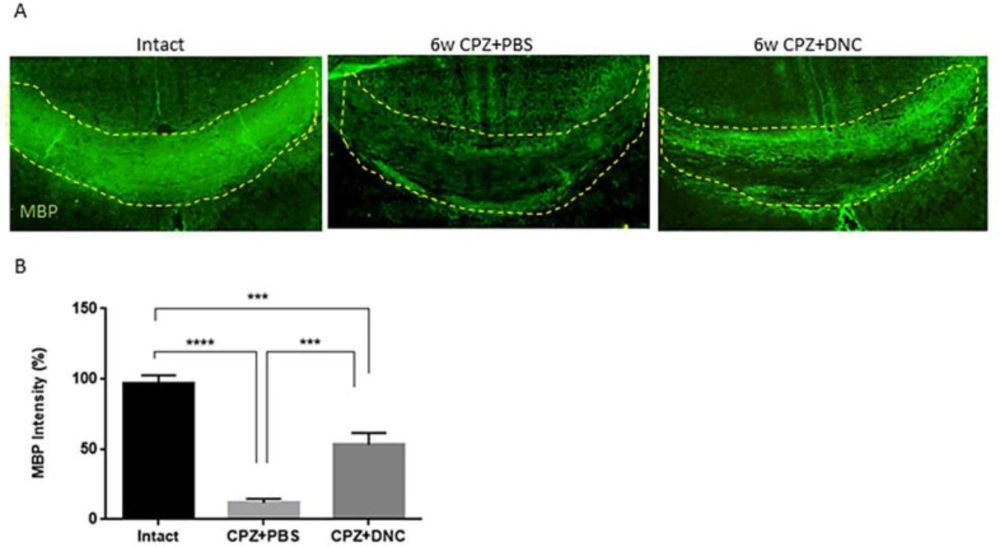
In order to further confirm the protective effect of DNC on myelin content of CC against CPZ, along with LFB, immunostaining for MBP as a marker of myelination was performed. Our findings showed that DNC treatment lead to higher degree of myelin intensity CC, compared to CPZ+PBS group (
Figure 4).
Several lines of evidences have indicated that OLs death and disruption of myelin integrity are associated with many pathological and neurodegenerative symptoms of MS. Thus, protecting myelin or myelinating cells at early stages of MS is likely to be of clinical relevance. Current therapies for MS are designed to modulate or suppress the inflammatory processes of the disease, but provide limited benefit for survival of myelinating cells and preservation of myelin sheaths. Even though, a number of studies has shown the anti-inflammatory, antioxidant, immunomodulatory, anti-astrogliosis properties of curcumin in MS (
39,
40), here we indicated a new feature of curcumin, as a nano-formulation, that protects myelin forming cells from apoptotic death by CPZ induced oxidative stress, apparently due to inhibition of inflammatory, non-permissive microenvironment induced by microglia and astrocytes. Although the potential underlying molecular mechanisms of these effects by DNC remains to be elucidated, the results from our study suggest DNC as a promising compound to protect or promote myelin index in context of demyelinating conditions. However, future studies are required to confirm the real benefits of DNC for the management of demyelinating diseases.
| Target | source | Labled cells | Company |
|---|
| Olig2 | Rabbit | Oligodendroglial lineage cells | Abcam, ab9610 |
| GFAP | Rabbit | Reactive astrocytes | DAKO, Z0334 |
| Iba1 | Rabbit | Microglia | sc-32725 |
| MBP | Mouse | Mylin | Abcam, ab40390 |
DNC treatment exhibited protective effect on OLLCs. A. A schematic image of experimental design. B) In-vivo imaging results showing that DNC is able to pass through intact BBB of normal mice (right). Under the same imaging conditions the level of fluorescence intensity in control mice was not indicated C) Representative image of brain section IHF by OliG2, maker of OLLCs. D) Analyzing the number of Olig2+ cells in intact, CPZ + PBS and CPZ + DNC showed that the number of OLLCs was significantly lower in PBC treated group compared to intact and DNC treated mice. However, no significant differebce was observed in the number of OLLCs in DNC treated group. n = 27 brain sections from 3 mice, per experiment group. Values are given as mean ± SEM as the results of ordinary one-way ANOVA followed by Tukey’s multiple comparison test
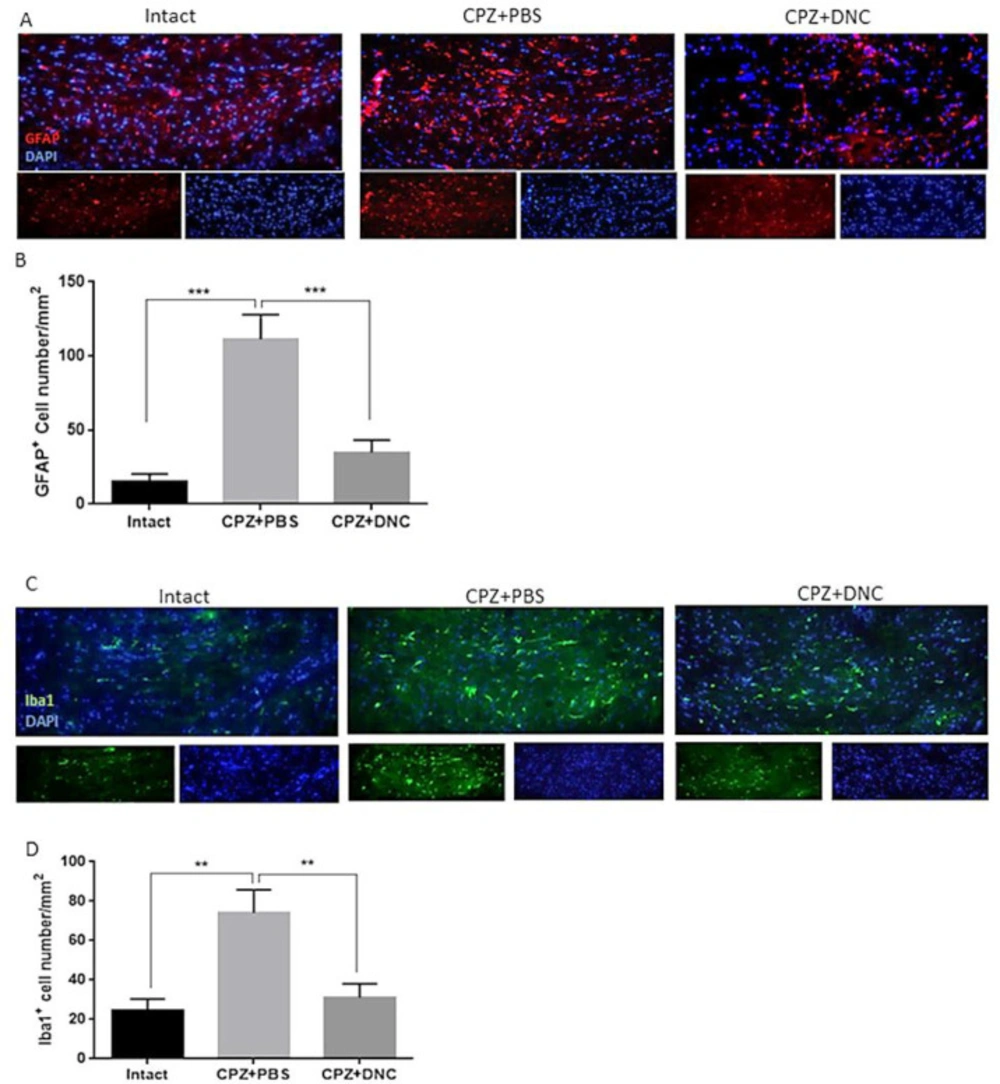
DNC treatment suppressed accumulation and activation of astrocytes and microglia in CC of CPZ treated mice. (A) Representative image of IHF of brain sections for GFAP, marker of reactive astrocytes. (B) Comparing the number of GFAP+ cells in intact group with CPZ + PBS group showed that CPZ treatment lead to significant increase in number of reactive astrocytes in CC; however, DNC treatment along with CPZ feeding, lead to significantly lower number of GFAP+ cells in CC. (C) Representative image of IHF of brain sections for Iba1, marker of microglia. (D) Comparing the number of Iba1+ cells between intact mice and CPZ + PBS group showed that CPZ intake lead to significant accumulation of microglia in CC; however, significant decrease in Iba1+ cells in DNC treated group showed suppressive effect of this compound on microglia activation by CPZ. N = 27 brain sections from 3 mice, per experiment group. Values are given as mean ± SEM as the results of ordinary one-way ANOVA followed by Tukey’s multiple comparison test
DNC treatment showed protective effect on myelin content of CC during CPZ intake. (A) Representative images for LFB staining results.( B) Evaluation of myelination index by analyzing LFB intensity indicated that even though in intact mice the myelin sheets showed normal, multi-layered and compact structure 6 weeks treatment with CPZ and PBS lead to disorganized myelin composition with significantly lower intensity, compared to intact mice. But, in group that received DNC along with CPZ intake the degree of myelination was significantly higher than PBS treated group. The degree of myelin, however, was still lower in DNC group, compared to intact mice. n = 27 brain sections from 3 mice, per experiment group. Values are given as mean ± SEM as the results of ordinary one-way ANOVA followed by Tukey’s multiple comparison test
DNC treatment showed protective effect on MBP content of CC after CPZ intake. A) Representative images from brain sections IHF for MBP, marker of main components of myelin sheath. B) Analysing the intensity of MBP in CC of mice from intact and CPZ+PBS group showed that 6 weeks CPZ intake lead to significant depletion in myelin content of CC; however, the degree of myelination was significantly higher in DNC treated group compared to PBS treated one. n = 27 brain sections from 3 mice, per experiment group. Values are given as mean ± SEM as the results of ordinary one-way ANOVA followed by Tukey’s multiple comparison test