Preparation of scFv phagemid library
scFv phagemid library was achieved from Tomlinson I (Life Sciences, Cambridge, UK) and transformed into E. coli TG1 bacteria. E. coli TG1 was cultured in a 30 mL SB medium (Super broth: 5 gr Yeast extract, 10 gr Tryptone and 10 gr NaCl for 1 liter) containing 100 µg/mL ampicillin. For emancipation of phage particles expressing scFv antibodies, when the OD600 reached 0.6, 1012 pfu M13K07 helper phage (Amersham Pharmacia Biotech, USA) was added and the culture was incubated in 37 °C for 30 min followed by another 30 min at 37 °C/250 rpm. Then, kanamycin was added at the final concentration of 70 µg/mL and the culture was incubated for 16 h at 37 °C/250 rpm. For separating the recombinant phage particles, the culture was centrifuged at 5000 rpm for 20 min and the supernatant was collected. Then 20% (v/v) PEG solution (polyethylene glycol 6000 in 2.5 M NaCl) was added to the supernatant and incubated for 4 h on ice. In order to eliminate the bacteria cells from supernatant, the supernatant was incubated in water bath at 60 °C for 20 min and bacterial debris were removed by centrifugation at 5000 rpm for 5 min. The solution was centrifuged at 4 °C /12,000 rpm for 20 min and pellet containing phage particles was resuspended in TBS (50 mM Tris–Cl, 150 mM NaCl, pH 7.5) solution containing 1% BSA. Finally, the supernatant was stored at 4 °C until the next use.
Bio-panning and polyclonal phage-ELISA
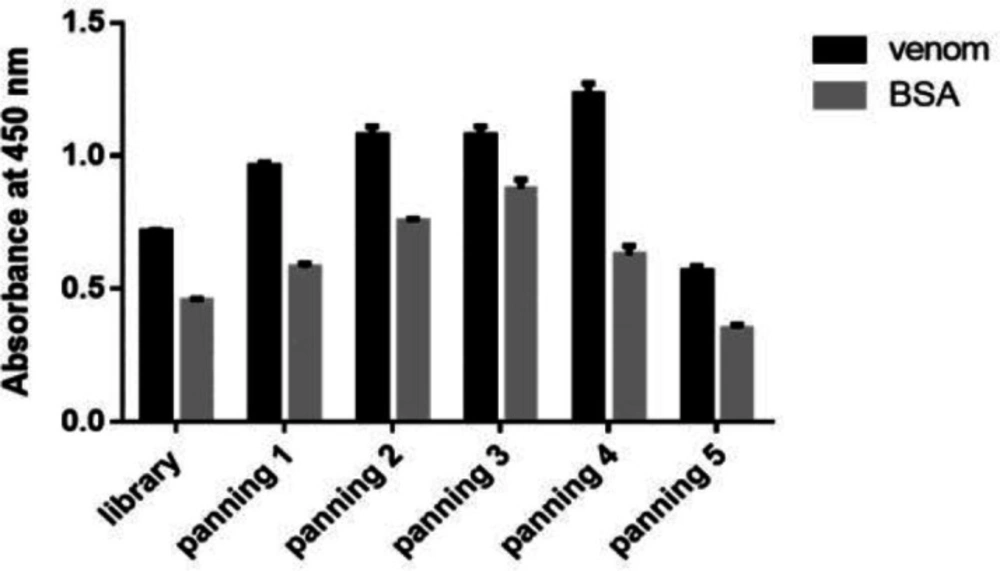
For the first round, 10 μg of the venom and 10 μg BSA were coated in microplate in duplicate and incubated for 16 h at 4 °C. The wells were washed three times with TBST 0.05% (TBS + 0.05% Tween-20) solution and blocked with 200 µL TBS buffer containing 3% BSA, then incubated for 2 h at 37 °C. After washing, 100 µL of recombinant phage library was added to wells containing BSA and incubated for 1 h at a temperature of 37 °C. Then the recombinant phage library solution from wells containing BSA was collected and added to wells containing the toxin and incubated for 1 h at 37 °C. In the next step, the wells were washed several times with TBST and the conjugated phages were eluted with 100 µL of Glycine-HCl (pH = 2.2). After 10 min, the released phages were neutralized with 40 µL of Tris buffer (pH = 9.2) and added to 30 mL of TG1 bacteria culture with OD600 of 0.6. The phages were propagated as described under “Preparation of scFv phagemid library”. The separated phages were used for the next round of panning. Five rounds of bio-panning were carried out with the toxin concentrations of 10, 5, 2.5, and 1.25 µg. In order to perform polyclonal phage ELISA, each well of the microtitre plate was coated with 10 µg of venom and BSA (as a non-specific antigen) and incubated at 4 °C for 16 h. The wells were washed and blocked as described before. Separated phages from each round of panning were added to corresponding wells and incubated at 37 °C for 2 h. After washing using TBST 0.05%, diluted anti-M13 monoclonal antibody conjugated to HRP (Amersham, Germany) was added to the wells and incubated at 37 °C for 2 h. the wells were washed three times using TBST 0.05% and TMB (3, 3′, 5, 5′-Tetramethylbenzidin) substrate (Sigma, USA) was added to the wells. After 20 min the reaction stopped with 3 N H2SO4 solution. Finally, the optical density was read at 450 nm.
Expression and purification of selected polyclonal scFv
Phagemids were extracted by plasmid extraction kit (Bioneer, South Korea) and transformed into HB2151 bacteria. The bacteria were grown in LB (Luria Bertani) medium until the culture reached an OD
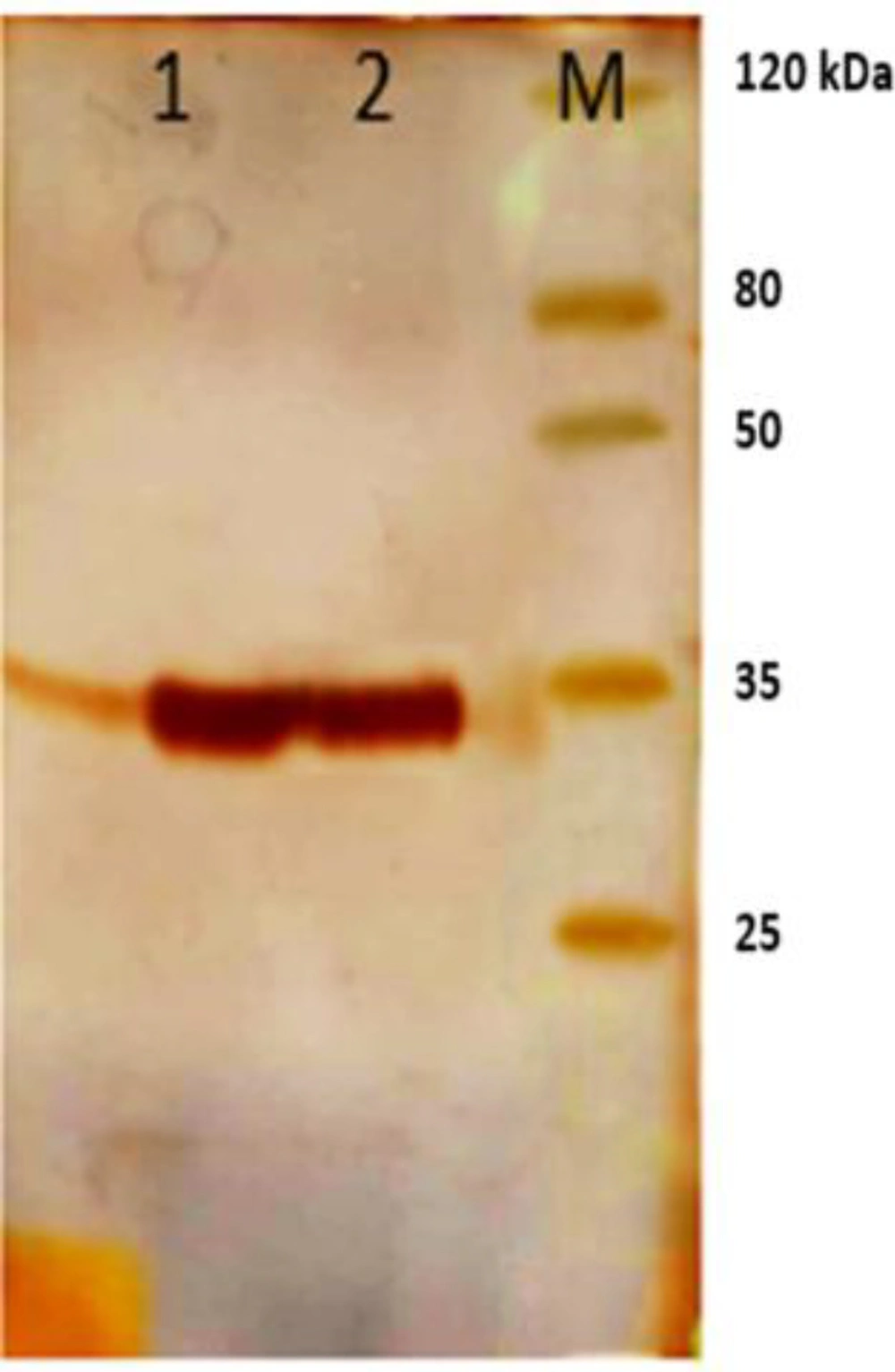
600 of 0.8. Expression of the polyclonal scFv was optimized with 1mM IPTG at 37 °C for 5 h. the bacterial cells were collected by centrifugation at 5000 rpm for 20 min and after adding 6 M urea solution to pellet, periplasmic proteins were extracted by sonication. Purification of scFv antibodies was performed using Ni-NTA chromatography (Qiagen, Germany). Results of the expression and purification were surveyed on 12% SDS PAGE (
Figure2).
Rat immunization and serum semi purification
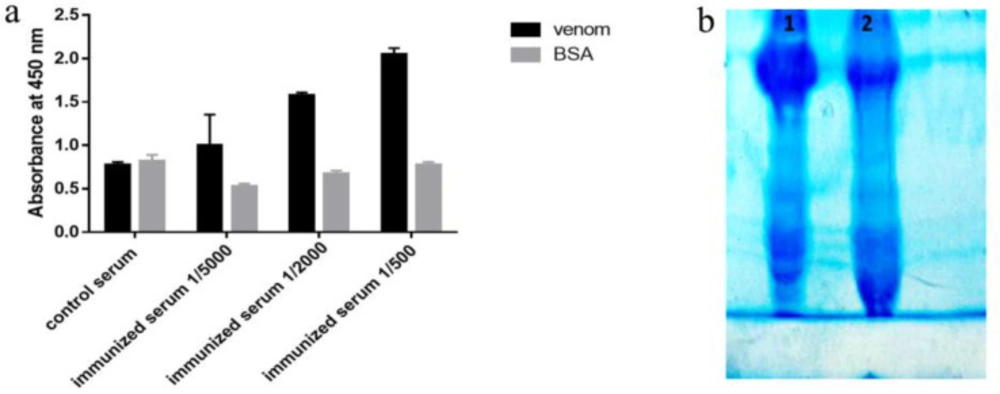
3-month-old rats were used for immunization. At first 200 µL of 250 µg/mL toxin solution was combined with 200 µL of Freund’s complete adjuvant and was injected intraperitoneally (IP). Subsequent immunizations were performed by a mixture of 200 µL of venom (at the concentration of 250 µg/mL) and 200 µL of incomplete Freund’s adjuvant, injected on days 10, 22, and 36 after first injection, respectively. To evaluate rat immunization, seven days after the fourth injection, 200 µL of bloods were collected and the antibody’s titer was determined using ELISA. Then the bloods from the control and immunized rats were collected and fractionated with ammonium sulfate. The amount of ammonium sulfate was calculated by the ammonium sulfate calculator software (http://www.encorbio.com). The results of semi-purification and immunization were analyzed on 7% SDS PAGE and ELISA, respectively. 96-Wells microtiter plate was coated with either 1.25 µg of venom or BSA in 100 µL PBS and incubated at 4 °C for 16 h. Then the wells were washed and blocked as previously was described. Dilutions of 1/500, 1/2000, and 1/5000 of immune sera and 1/300 of non-immune sera were prepared with PBS and added to the wells and were incubated at 37 °C for 1 h. The wells were washed with TBST 0.05% and then incubated with HRP conjugated anti-rat antibody (1/20000 in PBS) (Abcam, UK) for 1h at 37 °C. Following the final washing step, TMB was added to each well and after 20 min, the reaction was stopped with 3N H2SO4 solution. The optical density of each reaction was measured at 450 nm.
Evaluation of antibody binding
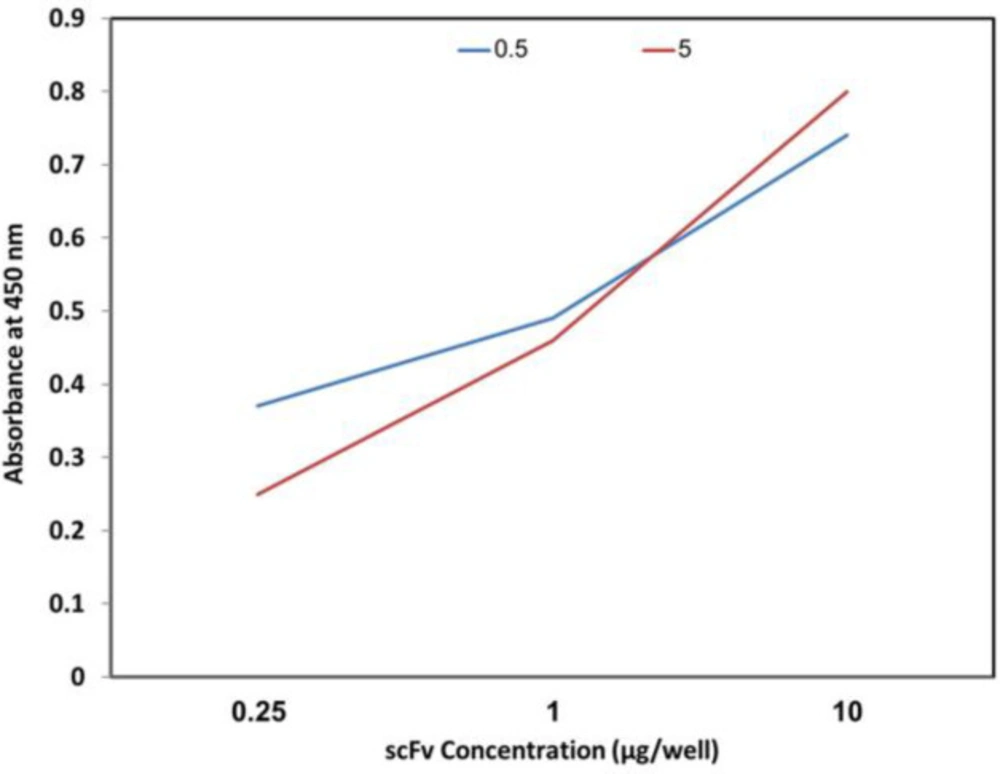
Venom in concentrations of 0.25, 1, and 10 µg per 100 µL PBS was coated in a 96-well microtiter plate in duplicate and incubated at 4 °C for 16 h. The following steps were carried out as described for polyclonal phage-ELISA except that purified scFv in concentration of 5 µg per 100 µL of PBS was used instead of phage, and anti-His antibodies conjugated with HRP (Abcam, UK) were applied instead of anti-M13 antibodies conjugated with HRP.
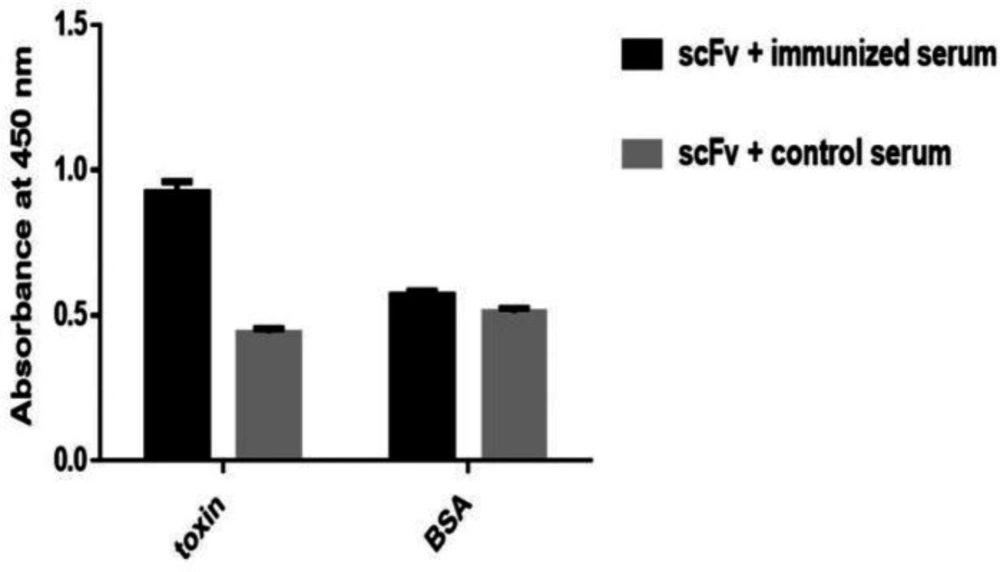
Evaluation Specificity of polyclonal scFv by Sandwich ELISA
The 96-Wells microtiter plate was coated in duplicate with the scFv as the capture antibody at a concentration of 5 μg/mL in PBS and was incubated at 4 °C for 16 h. Then wells were washed and blocked as previously described for polyclonal phage-ELISA (
21). 100 µL of venom and BSA in concentration of 5 μg/mL in PBS were added to the corresponding wells. The wells were washed and subsequently incubated with 1/1000 and 1/300 dilutions of immune and non-immune sera, respectively, and were incubated at 37 °C for 1 h. The rest of the steps were carried out as previously described under rat immunization and semi-purification serum.
Evaluation of LD50
The toxin was provided by Razi Institute. Dilutions for injection were prepared from the stock solution with 1.2 µg/µL concentration. BALB/c mice with 15-gr weight were divided to three groups and injected with 200 µL of toxin dilutions intraperitoneally in duplicates. The lethality was monitored 72 h post-injection and the toxin concentration in which 50% of the mice were survived was selected as LD50.
In-vivo Challenge of the Mice
First, lethal dose of venom (LD50) was determined. The LD50 of the venom was indicated 8.5 µg in BALB/c mice weighing 15 gr. To appraise the neutralization capability of the purified scFv, different concentrations of scFv and venom were injected intraperitoneally. The mice were divided into six groups. Venoms in concentrations >1 LD50 and 2 LD50 were mixed with different concentrations of scFv and after incubation at 37 °C for 1 h, were injected into the mice. The survival of the mice was monitored after 72 h.