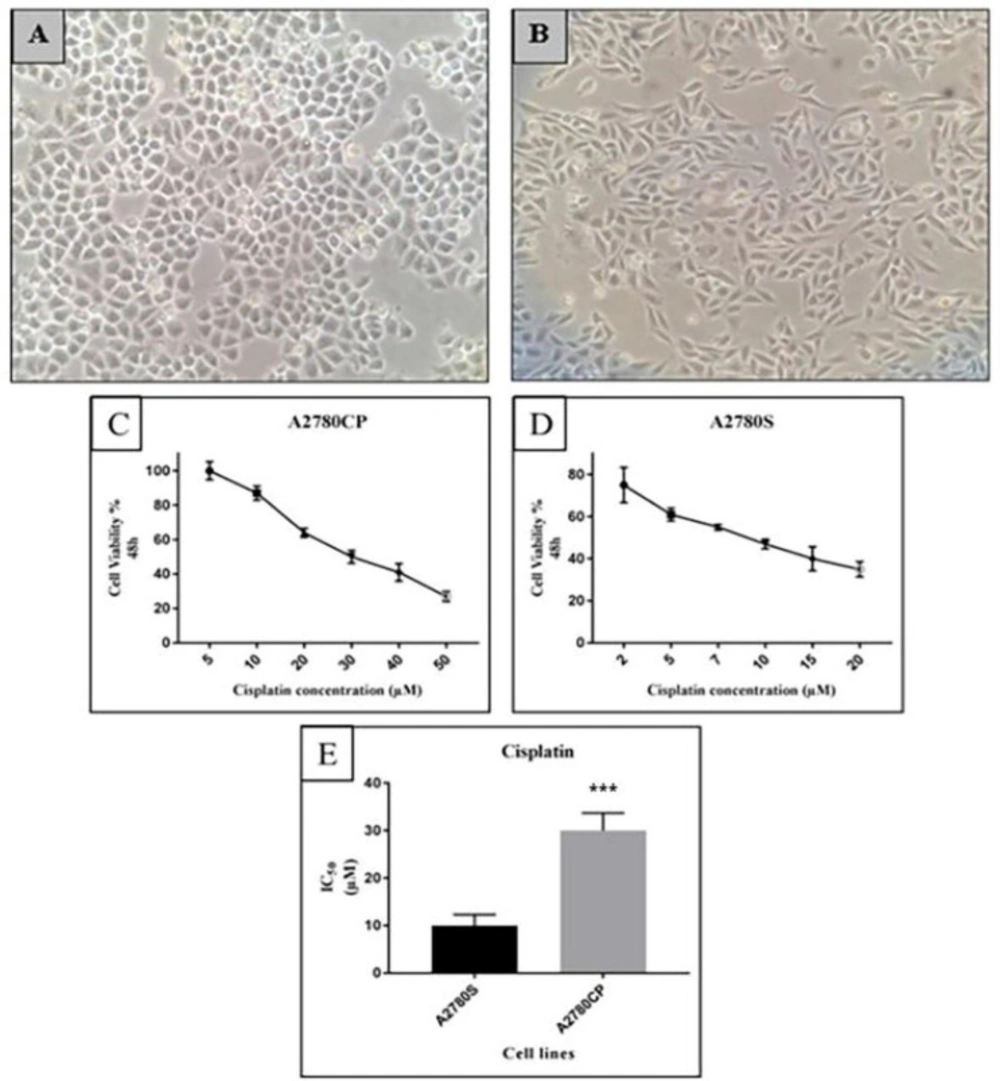
Cisplatin is the first-line chemotherapeutic agent for many malignancies including ovarian cancer. Cisplatin forms DNA adducts which inhibit DNA and RNA polymerases, interfere with DNA replication and transcription, and ultimately trigger apoptosis (
17). Ovarian cancer is the most common gynecological malignancy after breast cancer in women older than 40 years old particularly in developed countries (
18). One of the most promising strategies for overcoming chemoresistance in ovarian cancer is combining cisplatin therapy with other agents that enhance the anticancer effect of cisplatin (
14).
DNA hypermethylation and histone deacetylation have a major role in ovarian cancer chemo resistance, through silencing of drug resistance associated genes. Therefore, the inhibition of these epigenetic pathways can result in gene re-expression and consequently resensitization to chemotherapeutic agents (
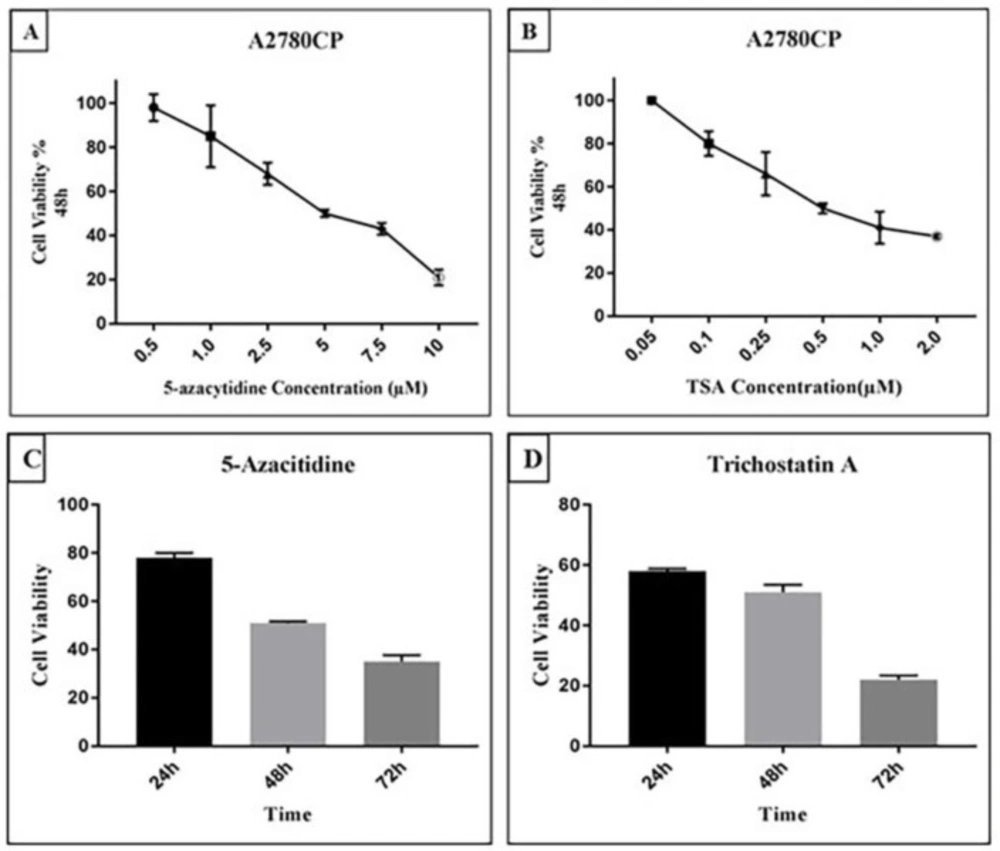
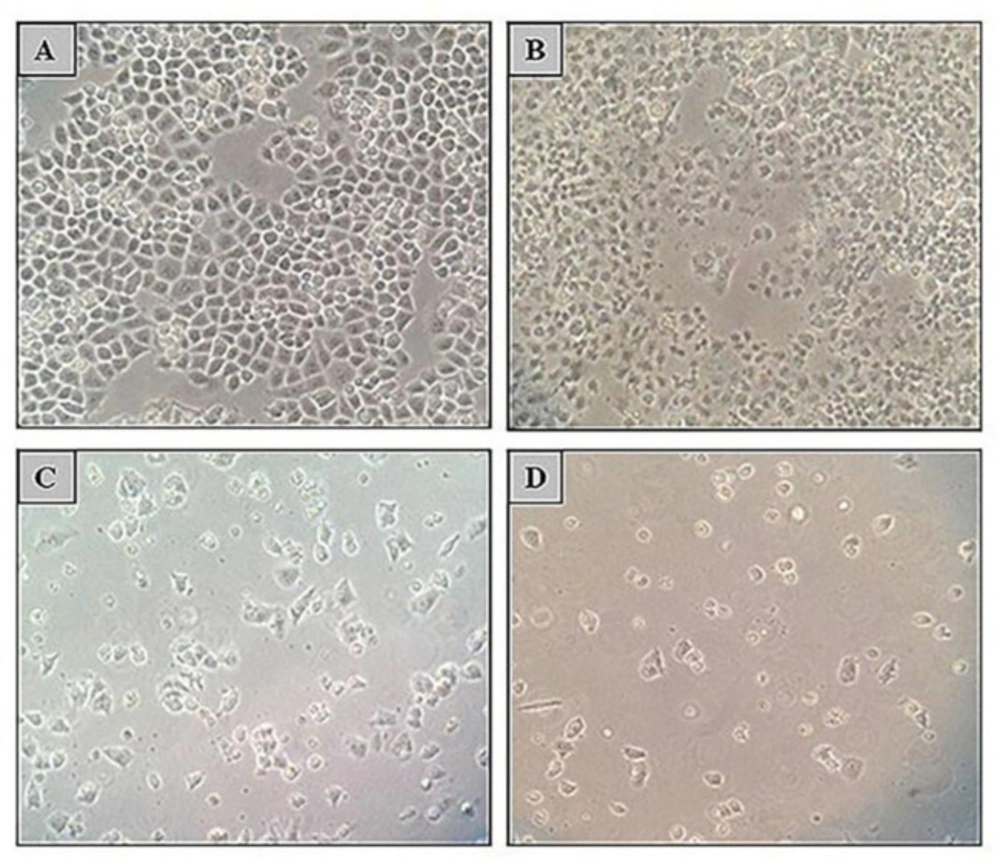
8). Our results revealed that 5-Aza and TSA can inhibit growth of ovarian cancer A2780 cells in a concentration and time dependent manner. 5-Azacytidine is a DNA methyltransferase inhibitor that was approved in 2004 by the Food and Drug Administration (FDA) for the treatment of myelodysplastic syndromes (MDS). 5-Aza can decrease the DNA methylation level and regulate the gene expression through reducing the bioactivity of DNA methyltransferase enzymes by trapping them for proteasomal degradation (
19). TSA is an antifungal antibiotic which inhibits class I and II HDAC enzymes by chelating zinc ion at the active site (
20). Previous studies indicated that histone deacetylation in hyper-methylated genes could result in gene inactivation while acetylation of histones could not lead to re-expression of silenced genes (
21). It has been claimed that TSA promotes Bax-dependent apoptosis in cisplatin-resistant ovarian cancer cells by up-regulation of p-73 (
22).
MiRNAs are small regulatory RNAs that are aberrantly expressed in different cancer types; (
23) and their role is demonstrated in the development of drug resistance (
24). A number of studies have asserted that overexpression of some specific miRNAs can enhance the sensitivity of ovarian cancer cells to chemotherapeutic agents. Chen
et al reported that transfection of miR-133b into paclitaxel and cisplatin resistant ovarian cancer cell lines results in increased sensitivity to paclitaxel and cisplatin (
25). In the other study, treatment with miR-142-5p mimic enhanced cisplatin-induced apoptosis in ovarian cancer cells (
26).
Many evidences show that miR148/152 family members act as tumor suppressor miRNAs and are down-regulated in many types of cancers as well as ovarian cancer (
13). MiR-148a targets various genes which affect a series of biological processes such as proliferation (IGF-IR, ROCK1, IRS1 and ERBB3), apoptosis (BCL-2), metastasis, and invasion (SMAD2, WNT1/10B, and USP4) (
27). Additionally, MET, TGFα, FGF2, CD151, and MMP3 are some of the oncogenic targets of miR-152 in human cancers; by targeting these genes, miR-152 leads to inhibition of cell proliferation and tumor metastasis (
12). It has been demonstrated that decreased expression of miR148a/152 is in association with increased tumor size and stages in gastric cancer (
28). It has also been observed that the lower expression of these miRNAs is inversely correlated with stage and lymph node status of breast cancer tumors (
29). Down-regulation of miR-148a and miR-152 leads to overexpression of DNMT1, promoting DNA methylation which could result in suppression of genes involved in chemo-resistance. Several genes silenced by promoter DNA hypermethylation such as MLH1, SULF1, SFRP, TNFRSF10A, UCHL1, and CLDN4 are related to platinum resistance in ovarian cancer. These genes are principally involved in DNA repair, apoptosis, cell growth, and invasion (
30). Restoration of these tumor suppressor genes could contribute to the re-sensitization of resistant ovarian cancer cells to platinum-based chemotherapeutic agents. Down-regulation of DNMT1 as a direct target of miR-148a and miR-152 by a negative feedback can result in higher expression of these miRNAs. Such a regulatory circuit has been reported in breast cancer; (
29) and might exist in ovarian cancer as well. In the present study, expression levels of miR-148a and miR-152 in A2780CP ovarian cancer cell line was investigated after treatment with TSA and 5-Aza using Real-Time PCR. We hypothesized that up-regulation of miR-148a and miR-152 by treatment with 5-Aza and TSA could reverse cisplatin resistance through DNMT1 suppression in ovarian cancer and miR-148a and miR-152 down-regulation might be due to DNA methylation and histone de-acetylation. DNMTs and HDACs strengthen the effect of each other in transcriptional suppression. Methylated DNA recruits HDACs either through methyl-CpG binding proteins or directly by DNMTs (
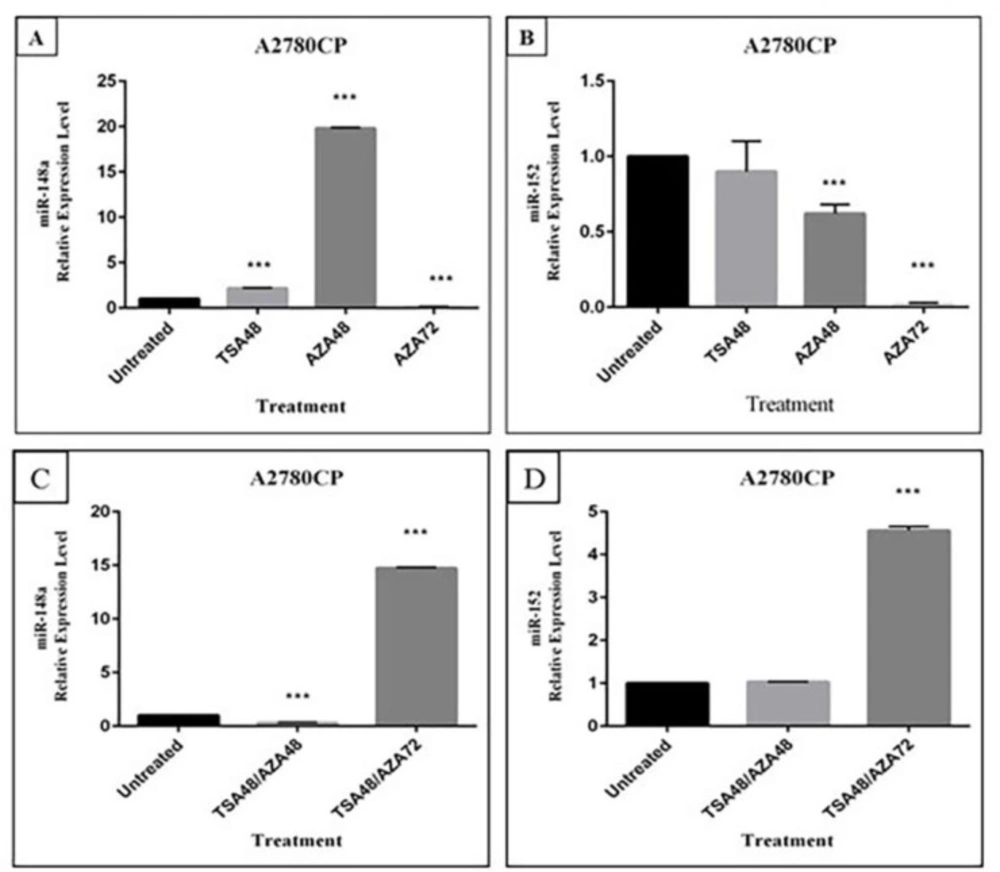
31). Therefore, combination use of DNMTi and HDACi may improve the limited anticancer efficacy observed with either therapeutic class alone. We observed that miR-148a was re-expressed in cisplatin-resistant cells when treated with 5-Aza and TSA alone and in combination; while miR-152 was only re-expressed when treated with a combination of these two drugs in comparison with control group. Surprisingly, miR-148a up-regulation after treatment with TSA for 48 h and 5-Aza for 72 h in association, was lower than treatment with 5-Aza for 48 h alone. These data indicate the potential of 5-Aza and TSA to reverse the expression of miR-148a and miR-152. The study by Cacan
et al., consistent with our observations, demonstrated that 5-aza-CdR and TSA treatment both enhance RGS10 (an important regulator of chemoresistance in ovarian cancer) expression and cisplatin-mediated cell death significantly in A2780CP ovarian cancer cell line alone and in combination. Further investigation demonstrated the contribution of HDAC1 and DNMT1 to silencing of RGS10 during acquired chemo-resistance and revealed the importance of HDAC1 and DNMT1 inhibition as an assistant therapeutic approach to overcome ovarian cancer chemo-resistance (
32). In another study on endometrial cancer cell lines it was revealed that hypermethylated miR-137 was overexpressed after co-treatment with 5-aza-CdR and TSA, more effective than treatment with each epidrug individually (
33). Liu
et al ,found that TSA and TSA plus 5-Aza treatment could significantly up-regulate RhoB and mediate apoptosis of SKOV3 and A2780 ovarian cancer cells, while 5-Aza could not do it alone (
34). In a recent study it was reported that 5-aza-CdR and TSA can significantly downregulate DNMT1 and HDAC1 and upregulate p21, p27, and p57 genes expression in colon cancer SW480 cell line separately but their joint application was not investigated (
35).
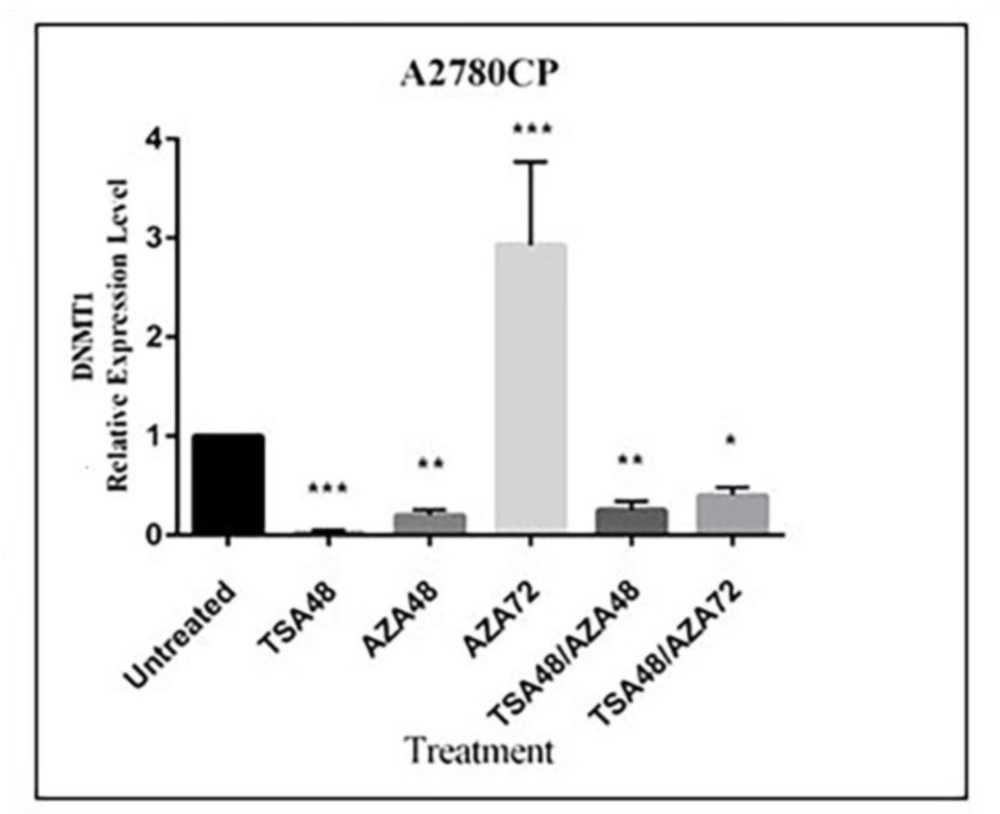
We demonstrated that miR-152 and miR-148a in cisplatin resistant A2780 cell lines were up-regulated after treatment with 5-Aza and TSA and consequently down-regulation of DNMT1 was observed. Even though DNMT1 is a direct target of miR-148a and miR-152, treatment-mediated up-regulation of the other hyper-methylated miRNAs targeting DNMT1 such as miR-185 on down-regulation of this gene should not be ignored. Xiang
et al ,revealed that over-expression of miR-152 and miR-185 leads to cisplatin sensitivity mainly through direct targeting of DNMT1(
14). We observed no significant up-regulation in miR-152 expression after treatment with DNMTi, 5-Aza. Our result is in consistent with their study that reported no significant change in miR-152 expression after treatment with 5-aza-CdR, a nucleoside analog inhibitor of DNA methyltransferases, in cisplatin-resistant A2780 and SKOV3 ovarian cancer cell lines. DNMT1 expression was interestingly down-regulated after 72 h treatment with 5-Aza; such an effect could be probably resulted from de-methylation of hyper-methylated transcription factors, after treatment with 5-Aza which consequently induces the expression of DNMT1. Examples of such hyper-methylated transcription factor genes are HOXA5; (
36) and members of the GATA family (
37). Findings of this study suggest an understanding of mechanisms by which epigenetic therapeutic options might provide a new approach to the treatment of ovarian chemo-resistance. In a recent study it was determined that combining DNMTi and HDAC6i upregulates expression of interferon stimulated genes and cytokines significantly more than each epigenetic drug individually in several cisplatin-resistant ovarian cancer cell lines. Thus it increases anti-tumor immune signaling from cancer cells (
38). Combined with platinum-based agents, DNA methyltransferase, and histone deacetylases inhibitors can sensitize resistant ovarian cancer cell lines to these chemotherapeutic agents and the same effect may be observed in animal models and the patients with recurrent ovarian cancer. Although the present study focused on miR-152 and miR-148a, some other microRNAs are likely to be involved in cisplatin resistance.