Chemicals
Curcumin (Sigma–Aldrich Inc., St Louis MO, USA), morphine (Temad Co., Iran), naloxone (Merck Co.), and dimethylsulfoxide (DMSO, Merck Co) were used in this study. The solutions were prepared by solving the chemicals in the normal saline freshly. Dimethylsulfoxide (DMSO) was used as the co-solvent to suspend curcumin in normal saline. A combination of normal saline and DMSO were used as the vehicle.
Animals
Male Wistar rats (weighing 250-300 g) were kept under optimum environmental conditions including the temperature of 22 ± 2 °C and a 12 h light-dark cycle. They had access to tap water and food freely. All the experiments of this study were carried out in accordance with the EU Directive 2010/63/EU for laboratory animals. The procedures which were implemented in this study were approved by the ethical committee of Shahid Beheshti University of Medical Sciences (# IR.SBMU.MSP.REC.1395.379).
Induction of physical dependence
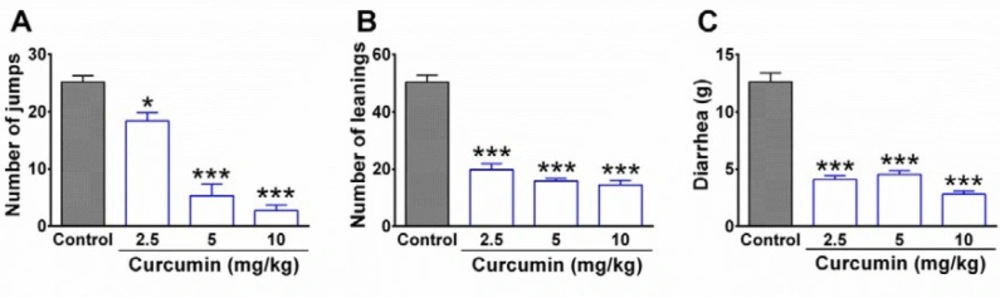
The animals were divided into five groups (n = 5 in each group; total numbers = 25) including control group, vehicle group, and three different curcumin-treated groups. In order to induce morphine dependence, all the animals in control and curcumin groups were given morphine sulfate intraperitoneally (i.p.) once daily for 9 days in an increasing dose of 10, 20, and 40 mg/kg, with each dose given for 3 days (25). The administration of morphine with a dosage of 40 mg/kg, i.p. was continued for a further 9 days. During the days 10th to 18th control group was given normal saline together with DMSO while curcumin-treated groups received three different doses of curcumin including 2.5, 5, and 10 mg/kg. In the vehicle group, the rats were served as intact animals and neither received morphine nor curcumin. Instead, the animals of the vehicle group were given normal saline (as the solvent of morphine) during the days 1st to 9th and received normal saline + DMSO 0.05% (as the vehicle of curcumin) between the days 10th and 18th. On the 19th day, six rats were selected randomly from each group for behavioral evaluation of withdrawal syndrome. The rest of the animals were considered for molecular experiments.
Evaluation of morphine withdrawal synd-rome
In order to evaluate physical dependence to morphine, the behavioral symptoms of withdrawal syndrome were evaluated in rats. A day after the end of the treatment period (day 19
th), six animals in each group were given naloxone (3 mg/kg, i.p.) as the opioid receptor antagonist. The rats were immediately placed in a transparent Plexiglas cylinder (30 cm in diameter and 50 cm height) and the behavioral symptoms of withdrawal syndrome including jumping, leaning, and diarrhea were evaluated in animals for 30 min (
16).
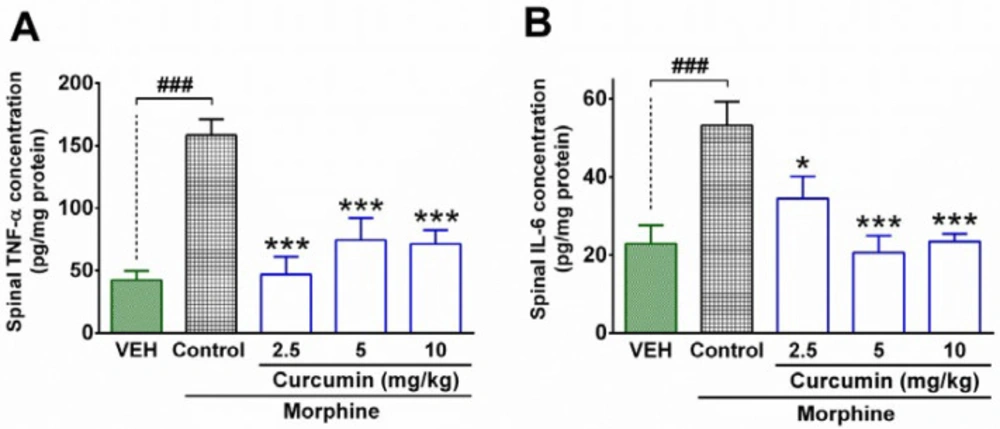
Measurement of inflammatory cytokines by ELISA test
In order to determine the spinal levels of inflammatory cytokines, the enzyme-linked immunosorbent assay (ELISA) method was utilized. At the end of treatment period (day 19
th), the other 4 animals in each group were served for measurement of inflammatory cytokines including IL-6 and TNF-α. First, the rats were sacrificed under anesthesia (ketamine 50 mg/kg, i.p., xylazine 5 mg/kg, i.p.) and the lumbar parts of the spinal cord of animals were removed immediately. Then, the concentration of protein in the spinal samples was determined by using the Bradford protein assay based on the method described previously. After the determination of protein contents in spinal samples, the concentration of IL-6 and TNF-α in the lumbar part of the spinal cord was measured by using a sandwich ELISA method. The procedure was conducted in accordance with the manufacturer’s instructions (ab100785 kit, Abcam and RAB 0312-1KT, sigma). An AQ8 ELISA reader (Biotech, USA) was used to determine the amount of absorbance in the standard solutions and spinal samples. The concentration (pg/g protein) of IL-6 and TNF-α in the sample was measured by using standard curves (
26).
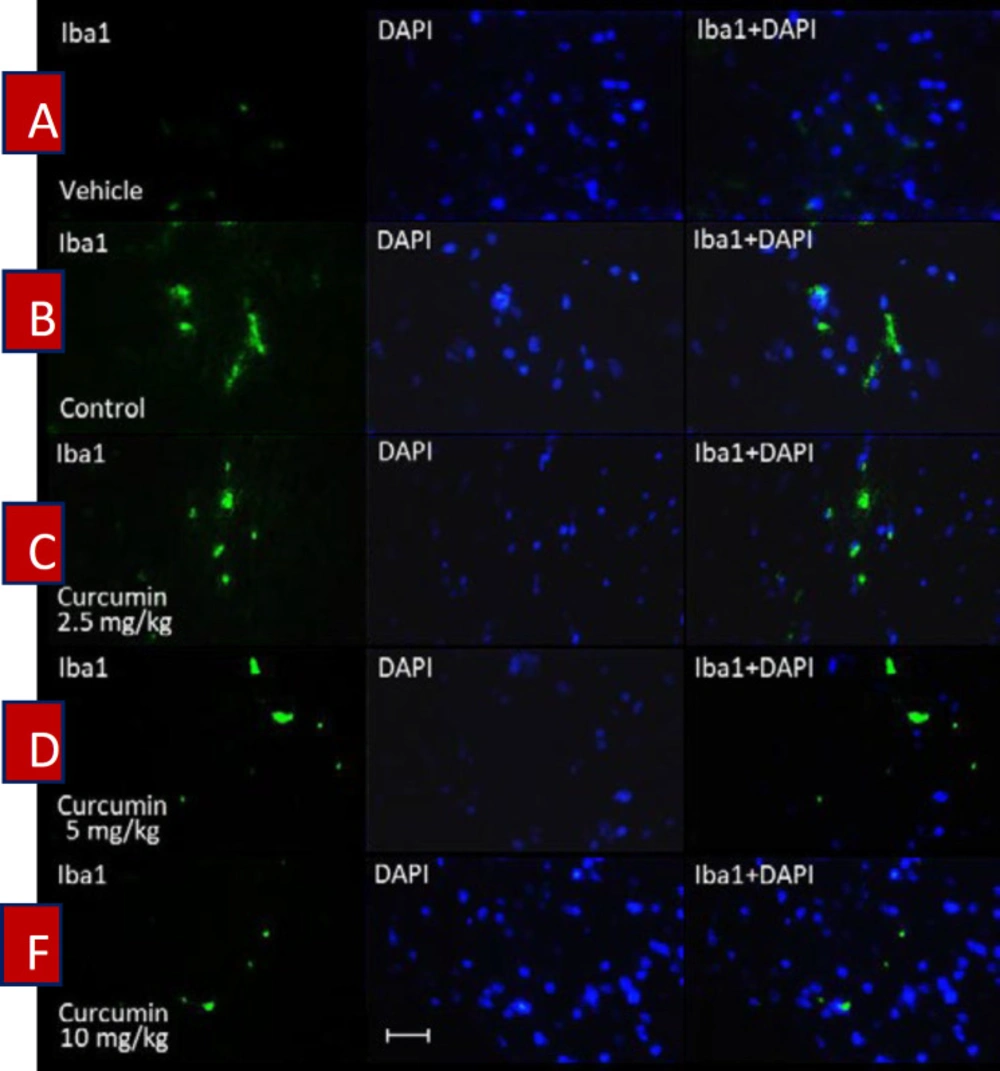
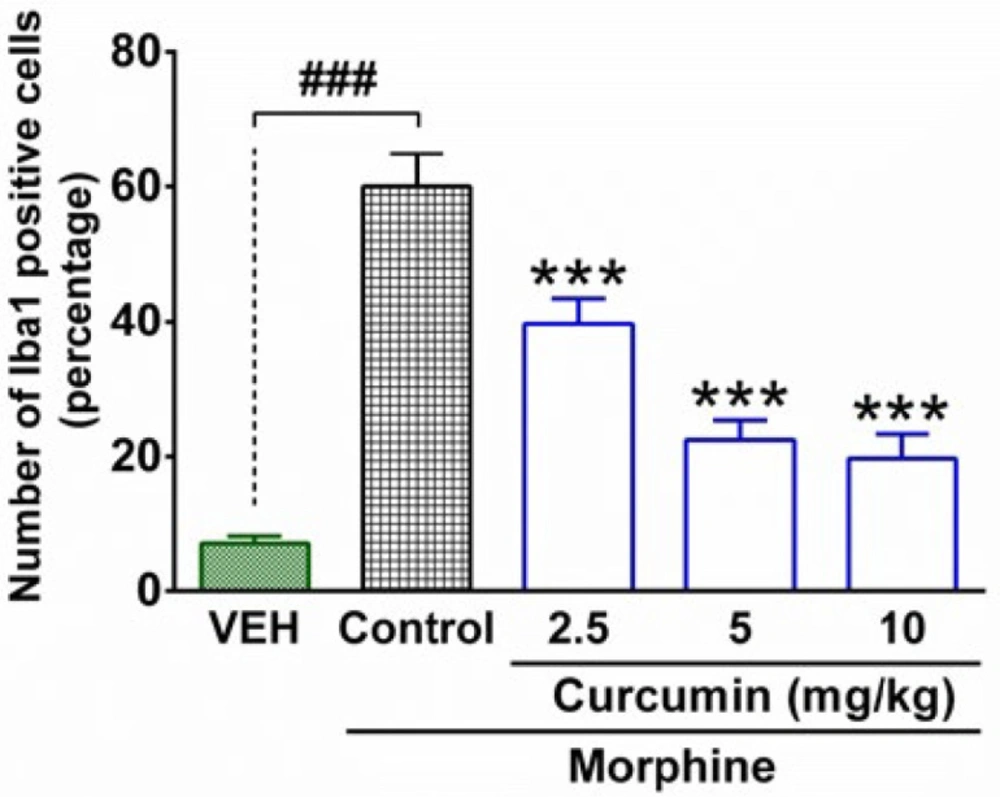
Spinal tissue preparation for Iba1 immuno-staining
Four remaining rats from each group were sacrificed under anesthesia at the end of the trial (19
th day). Then, the lumbar part of the spinal cord was dissected out from rats and transferred to formalin solution 4% in Phosphate-buffered saline (PBS). The spinal tissues were kept in the refrigerator (4 °C) for one week. Then, the samples were transferred to tissue processors for dewatering. After 16 hours, the samples were placed in tissue cassettes and were embedded in paraffin. Finally, the paraffin blocks of samples were cut coronally into 5 μm slices and were placed on the microscope’s slides. In order to determine the activation of microglial cells, the ionized calcium-binding adapter molecule 1 (Iba1) was stained by using the Anti-Iba1 antibody according to the manufacturer’s instructions (Abcam). Briefly, the samples were washed (2 × 5 min) in TBS (tris-buffered saline) plus 0.025% Triton X-100 with gentle agitation. Then, the slides were incubated in 10% normal saline with 1% BSA (bovine serum albumin) in TBS for 2 hours at room temperature. After that, the slides were drained for a few seconds and then wiped around with tissue paper. In the following, the samples were incubated with primary antibody overnight at 4°C. The slides were rinsed (2 × 5 min) with TBS plus 0.025% Triton X-100 and then incubated in 0.3% H
2O
2 in TBS for 15 min. Afterward, the samples were incubated with a secondary antibody for an hour at room temperature. At the end of staining, 4′,6-diamidino-2-phenylindole (DAPI) was used to identify the neurons’ nucleus. The slides were placed in DAPI for 20 min at room temperature and eventually washed with water for 5 minutes (
27). Finally, immunofluorescence microscopy was implemented to evaluate the intensity of immunostaining of microglial cells in the spinal samples. DAPI (4′,6-diamidino-2-phenylindole) staining was used to identify neurons’ nuclei.
Data analysis
The data were expressed as mean ± SEM for each group. Statistical analysis was carried out using one-way ANOVA, followed by Tukey’s test to compare the mean of each group with the mean of every other group. P < 0.05 was considered significant for statistical analysis.