Ethics statement
The experiments were performed based on the Principles of Laboratory Animal Care by the US National Institute of Health (NIH publication No. 85-23, revised 1985).
Cell culture
4T1 mouse mammary carcinoma cells were purchased from the Pasteur Institute of Iran (C604) and cultured in 75 cm2 culture flasks with Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum, penicillin (100 IU/mL) and streptomycin (100 mg/mL) until they were 80–90% confluent.
MTT assay
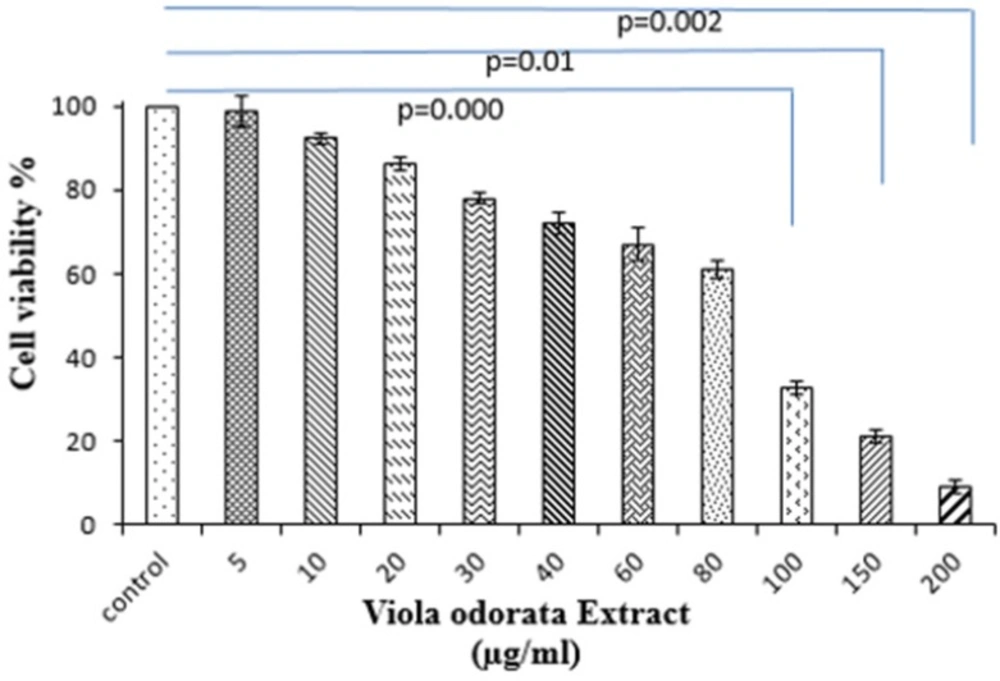
Cells were seeded at a density of 1×104 cells/mL in 96-well polystyrene culture plates at 37 °C with 5% (v/v) CO2 for 24 h. After incubation, cells were incubated with various concentrations of VOE (5, 10, 30, 40, 50, 60, 80,100,150 and 200 µg/mL). Control cells received only culture medium. Then, 100 µl of MTT solution (5 mg/mL) was added to each well and the plates were incubated at 37C for an additional 3 h. After dissolving the formazan crystals, cells were incubated with detergent at 37 °C. Absorbance was measured in each well, including the blanks at 570 nm by ELISA Plate Reader. Cell viability (%) was calculated for all groups compared to control sample. All experimental samples were performed in triplicate.
Viola odorata Hydro-alcoholic Extract
Aerial parts of
V. odorata were collected in summer from Rostamabad, Gilan, Iran and authenticated by Dr. Mohammad Reza Kanani. The plants were kept in the herbarium (Index Herbarium code: MPH-615) for future reference. Water-ethanol extract is a specific solvent for extracting antioxidants and polyphenolic compounds like flavonoids. In this study, the traditional solvent extraction (TSE) (50:50) method was used (
12). Department of Biology, Medicinal Plants and Drugs Research Institute (Shahid Beheshti University, Iran) prepared hydro-alcoholic (50:50) extract.
DPPH radical scavenging activity (DPPH assay)
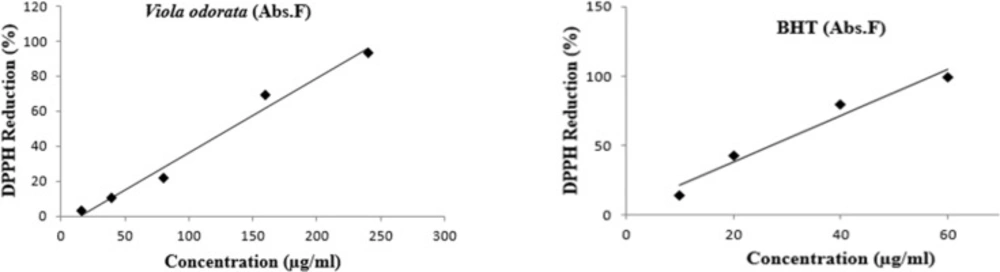
The scavenger activity of VOE was investigated by DPPH (α, α-diphenyl-β-picrylhydrazyl) assay. The solution of V. odorata (2 mg/mL) was prepared in methanol. The concentration of DPPH solution was 0.08 mg/mL in methanol. Synthetic antioxidant, Butylated hydroxytoluene (BHT) in 0.5 mg/mL concentration was used as a reference. VOE with different concentrations (2, 5, 10, 20, and 30 mg/mL) was transferred by pipette into columns of a 96-well micro plate. For methanol evaporating, plate was left at room temperature. In the first column, methanol (187 μl) and DPPH (63 μL) were added to four wells (control column) and 250 μL of methanol was added to remaining wells. The plate was shaken and allowed to stand in the dark place at room temperature for 70 min. Absorbance was measured in each well at 517 nm using plate reader. The following formula was used to calculate inhibition percentage (Percentage of free radical scavenging activity):
Ac is control absorbance, As is sample absorbance and Af is final absorbance. IC50 values are mean±SEM, n = 4. The lower IC50 in DPPH indicated a higher antioxidant activity.
HPLC of Viola odorata hydro-alcoholic extract
Different phytochemicals were isolated from all extracts i.e. n-hexane, butanol, methanol and aqueous extracts of
V. odorata like glycoside, saponins, methyl, salicylate, cyclotides, flavonoids, alkaloids, triterpenoids, mucilage and vitamin C (
7,
13-
15).
In-vivo and
in-vitro studies show that flavonoids have protective effects against many types of cancers(
16). Therefore, HPLC fingerprint was applied for qualification of the
Viola odorata extract and detecting the presence of flavonoids. HPLC protocol was used as described by Siddiqui et al. 2011 (
17).
V. odorata was extracted and dissolved in methanol at room temperature for 24 h. 10μL application volume was injected into HPLC and peaks with same retention time (RT) values with the standards were recorded.
Animal model
Tumor transplantation was carried out according to the modified method of
Pulaski et al, (2001) (
18). Female BALB/c mice (n = 20) weighing 18-20g at the age of 7-8 week were purchased from the Pasteur Institute, Karaj, Iran. 4T1 cells, cultured
in-vitro, were harvested and re-suspended serum free media. The animals received 0.1 mL subcutaneous injections of cell suspension (0.8 million cells) in the right mammary gland or hind flank. The animals were kept in animal house in standard condition and feed with standard pellet diet and water ad libitum, under controlled temperature (22 ± 0.5 °C) and light (12 h light/ dark cycle).
Experimental Protocol
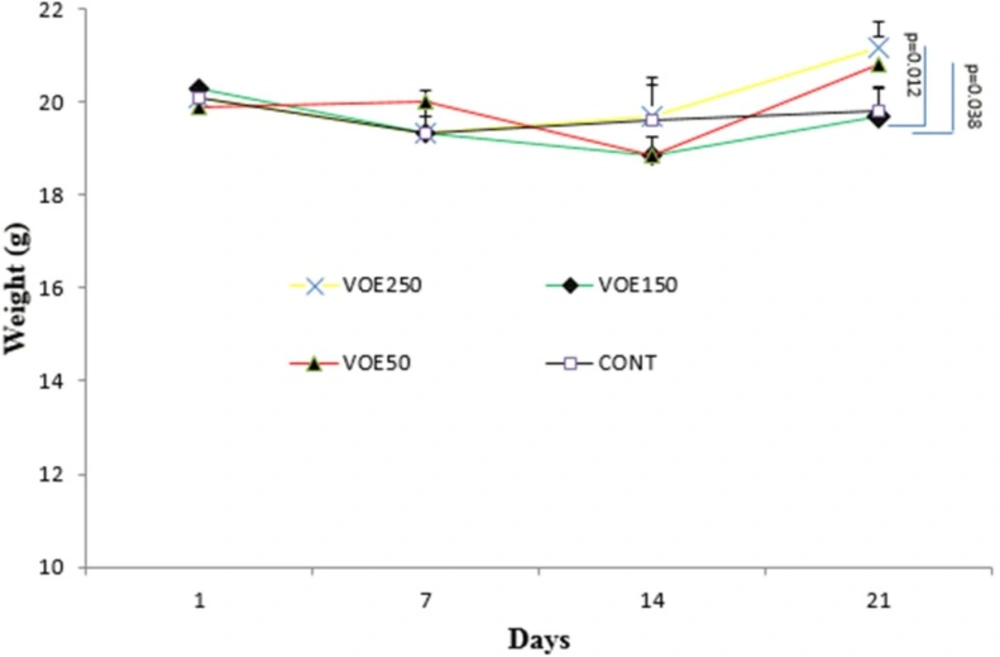
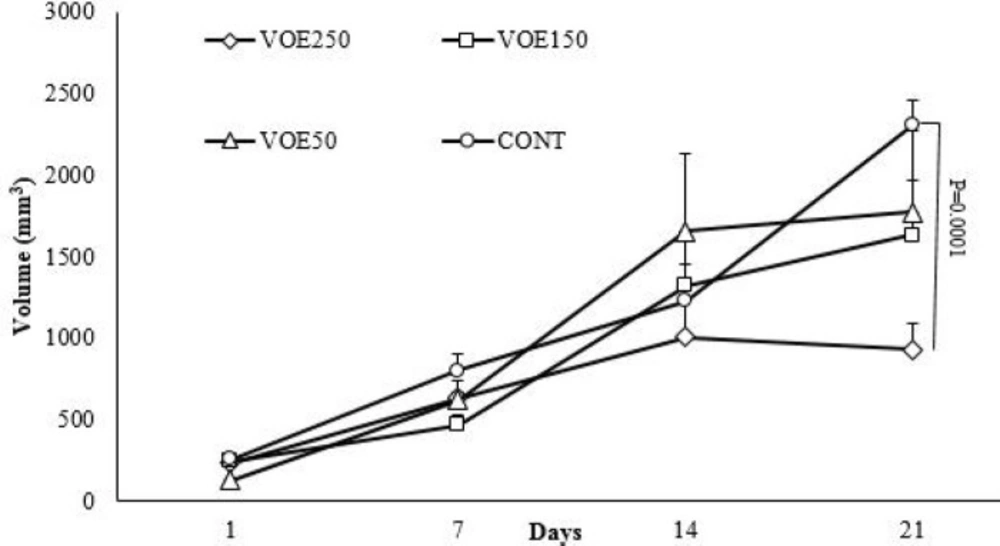
Animals were randomly divided into four groups (n=5). The first (VOE250), second (VOE150) and third (VOE50) groups were treated with 250, 150, and 50 mg/kg/day of the VOE for 3 weeks, respectively. The fourth group as the control group (Cont), received distilled water by gastric gavage. The mice were sacrificed on 22th day. The following formula was used to calculate the tumor inhibition rate and tumor volume:
Tumor inhibitory rate = (tumor weight of the control group − tumor weight of experimental group)/tumor weight of the control group ×100.
Tumor volume = ½ (length × width2)
Blood biochemical assays
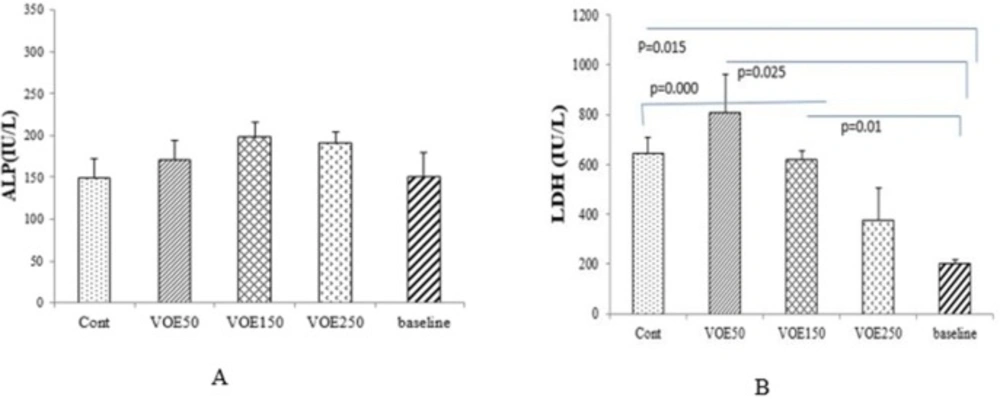
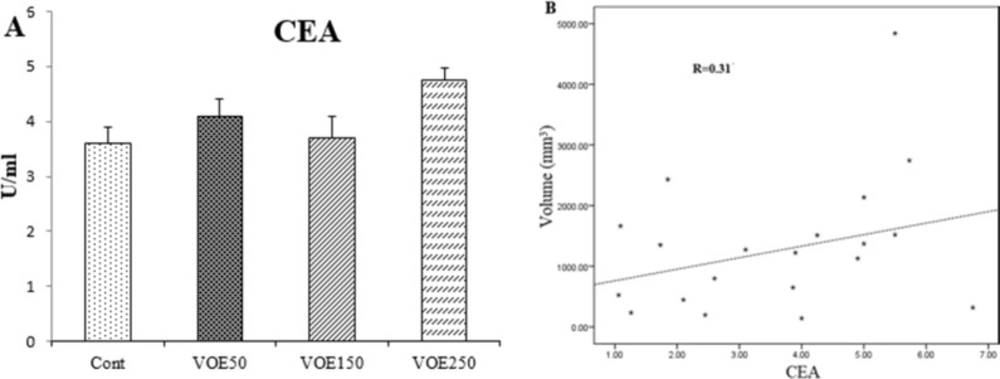
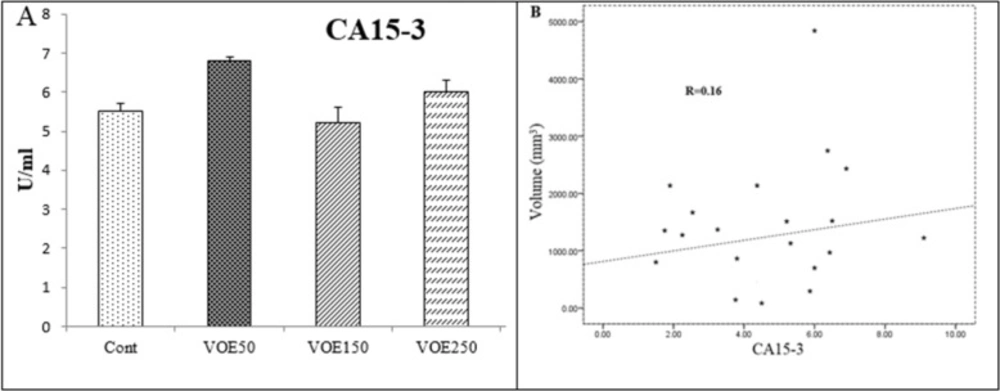
The blood samples were obtained from the heart of animals. In addition, 6 normal mice were used for obtaining baseline levels, on 22th day. The serum samples were collected after blood centrifugation at 3000 rpm for 10 min. These samples were used for determination of liver enzymes (GGT, ALP and LDH) and tumor markers (CEA and CA15-3) levels by means of commercial kits.
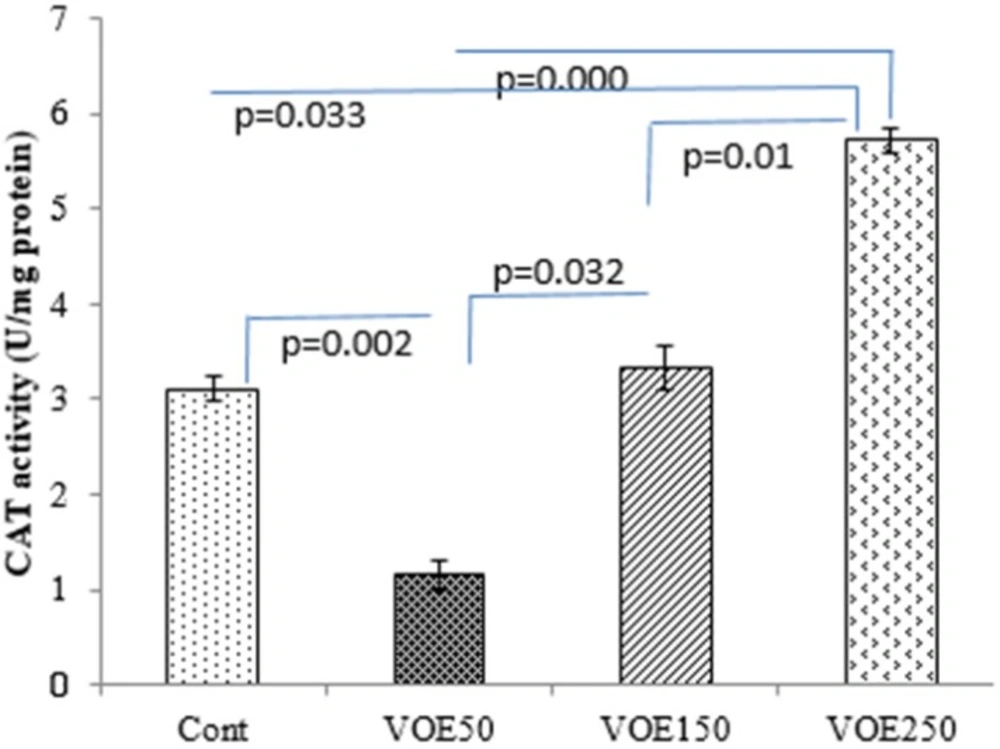
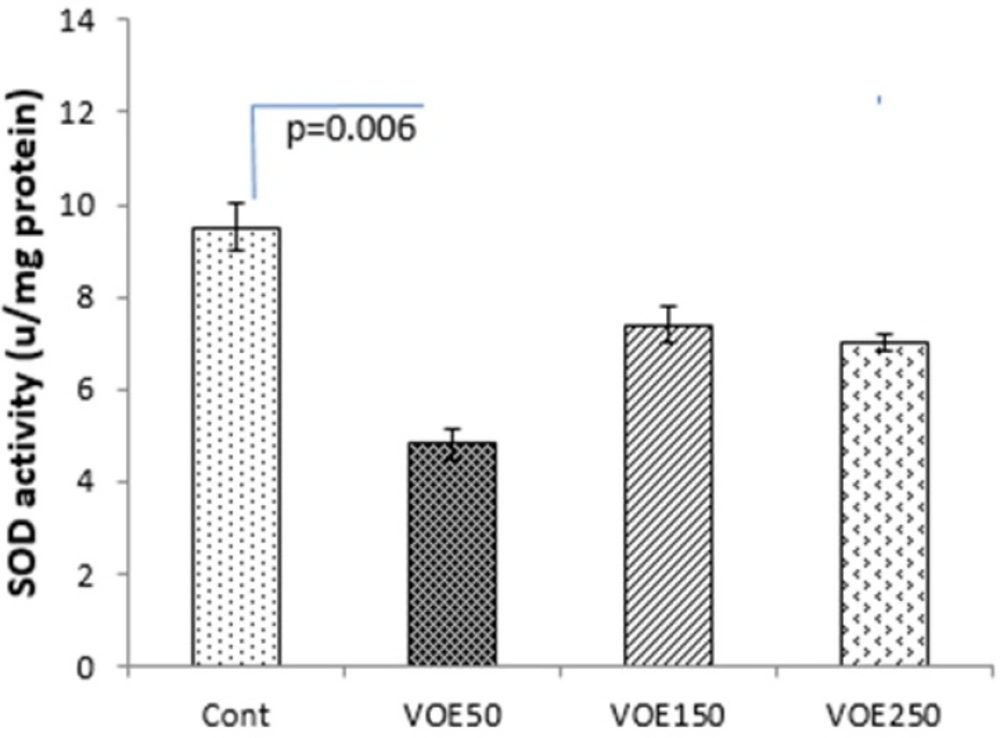
Determination of catalase and superoxide dismutase activities
CAT and SOD enzymes activities were assessed by the following method of
Genet et al., (2002) (
19) with some modifications. Catalase buffer was made by adding 0.05 mL H2O2 to 50 mL of sodium phosphate buffer and the rate of H
2O
2 decomposition was followed by monitoring absorption at 240 nm for 2 min. Superoxide dismutase buffer was made by adding 0.0018 gr EDTA and 0.003 gr pyrogallol to 50 mL of sodium phosphate buffer. The absorbance change at 420 nm of the mixture was monitored for 3 min.
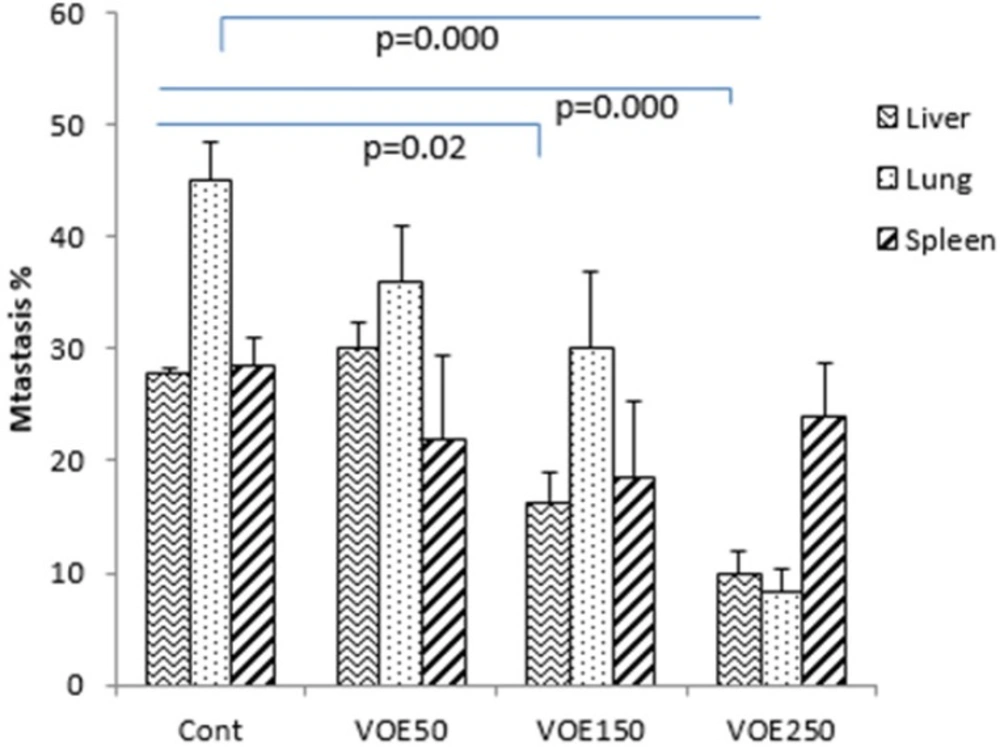
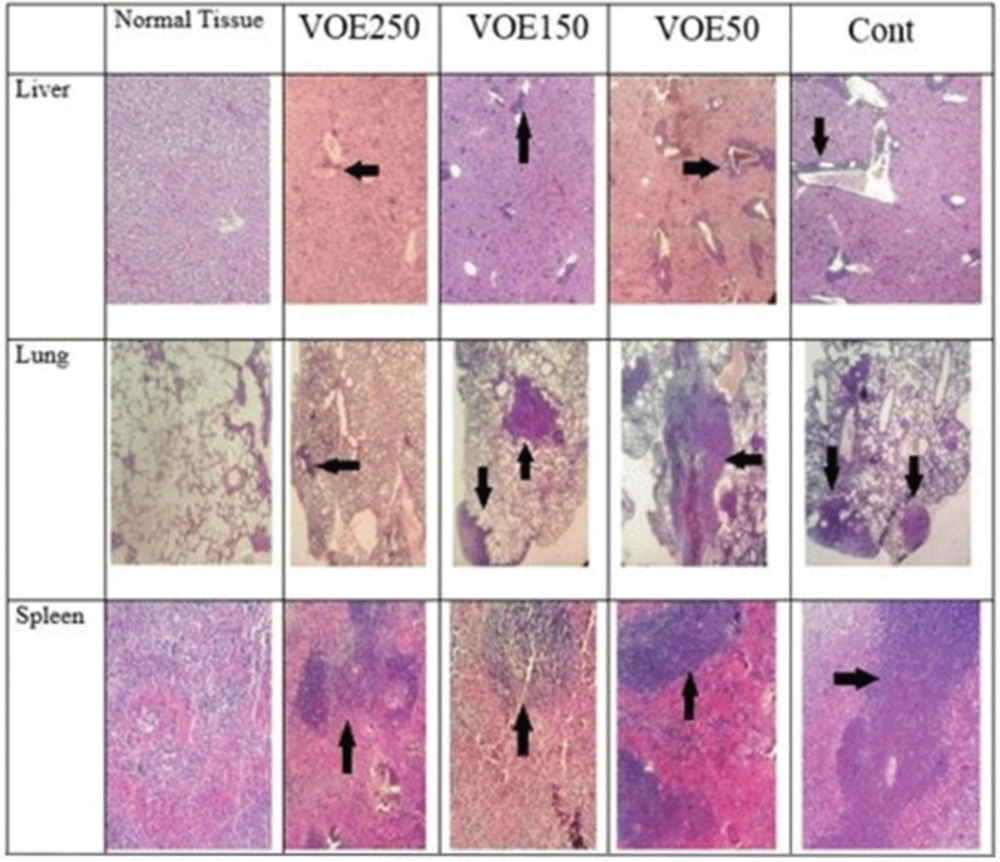
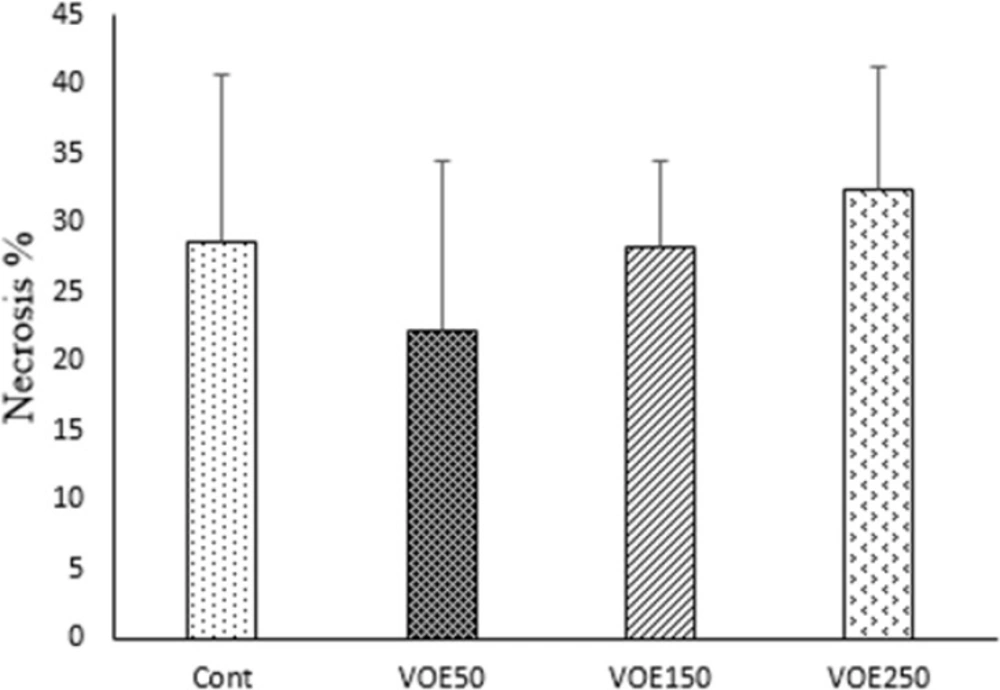
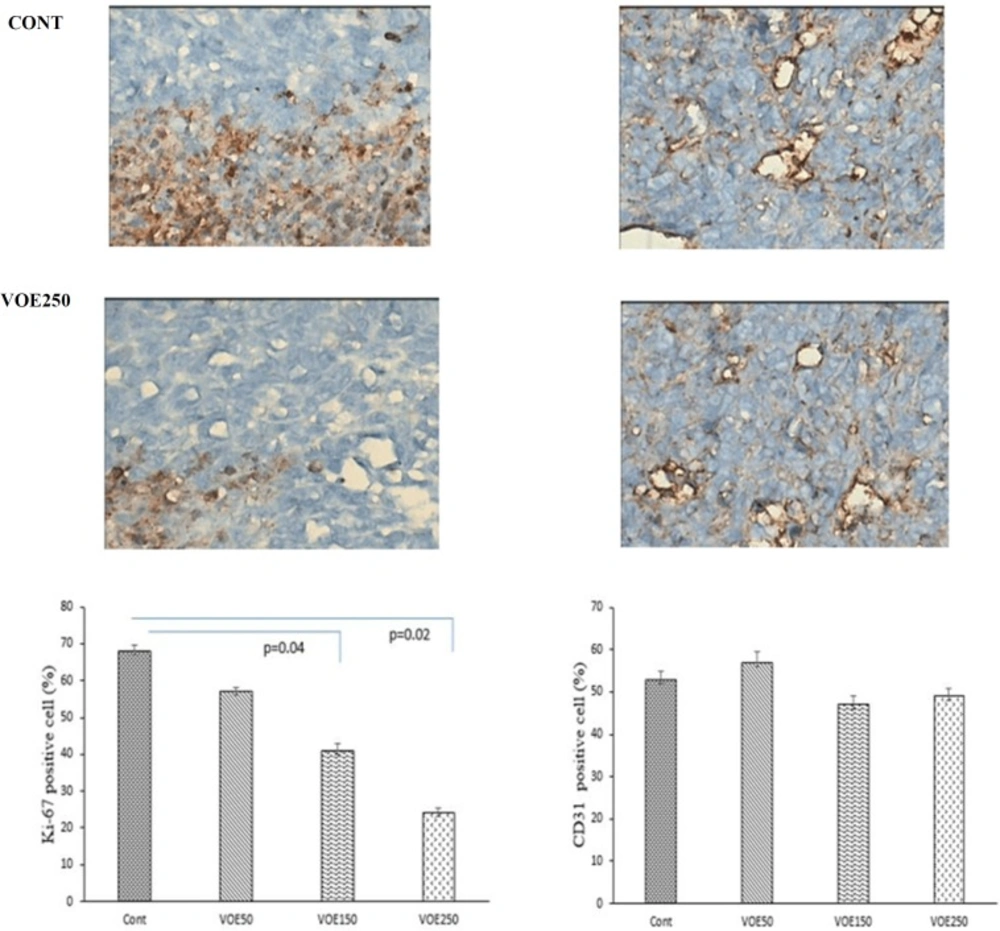
Immunohistochemical Analysis of Ki-67 and CD31 and histopathological studies
Liver, lung and spleen tissues were fixed, dehydrated and embedded in paraffin. Sections were stained with hematoxylin and eosin (H&E). Histopathological analysis was used based on H&E staining to identify tumor metastatic and necrotic area by ImageJ software as a percentage of total area. Tumor sections of control and VOE-treated mice were processed for immunohistochemical analysis of Ki-67 (Dako) expression to assess cell proliferation, and CD31expression (Dako) to
visualize blood vessels.
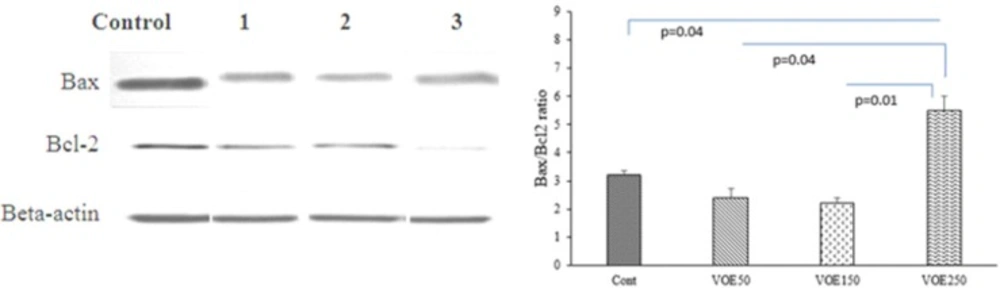
Western Blotting
The expression of Bax and Bcl-2 proteins were evaluated in the experimental groups (VOE250, 150, 50) and control group by Western blotting following sample extraction and SDS-PAGE. Breast cancer tissue samples were lysed in RIPA buffer (150 mM NaCl, 1% NP-40, 50 mM Tris PH=8.0, 1% SDS, 0.5% sodium deoxycholate, 1mM EDTA and protease inhibitor cocktail) and centrifuged at 12,000 rpm at 4 ºC for 20 min.
SDS sample buffer was added to aliquots of tissue extracts. Samples were heated at 100 ºC for 5 min. Protein was separated by 10% SDS-PAGE. Blots were incubated with specific primary polyclonal rabbit antibodies against Bax (1:500 dilution) (Santa Cruz), Bcl-2 (1:500 dilution) (Santa Cruz) and β-actin (1:1000 dilution) (Santa Cruz) in TBS-T for 18h. Then, they were incubated by secondary anti-rabbit (1:500 dilution) (Santa Cruz) in TBS-T for 90 min separately. Bax and Bcl-2 immune-reactive proteins were detected with advanced chemiluminescence (Enhanced Chemiluminescence, Amersham Biosciences) and exposed to a film. The signal intensity of the blots was measured by an image analysis system (Image j, version 1.46r).
Statistical Analysis
The obtained results were expressed as mean ± SD. Comparison with appropriate control was performed using one-way ANOVA. Correlation levels between tumor markers with tumor size and liver enzyme with metastatic rate were analyzed by a Pearson correlation coefficient (R). The least significant difference (LSD) was used as the post-hoc test. P < 0.05 was considered significant. Data was analyzed using the statistical package “SPSS 16.0”.