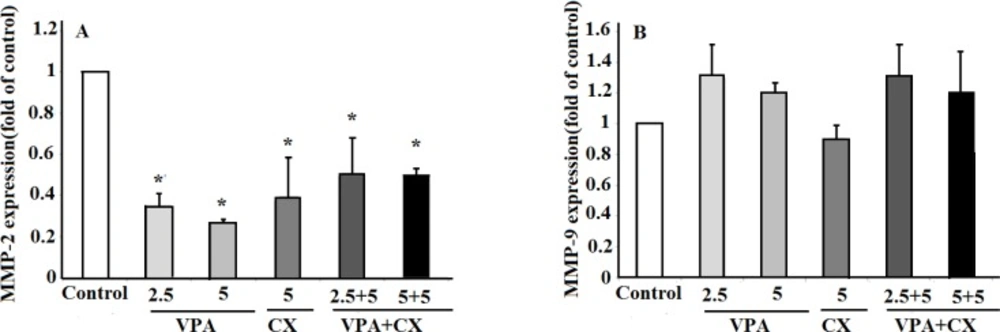
Due to role of metastasis in the poor outcome and mortality rate of PTC patients, finding of an efficient therapeutic modality that strongly inhibits growth and invasiveness characteristics of PTC cells is necessary. Combination therapy could be beneficial in this issue. In the present study, we evaluated the synergistic effects of CX and VPA on cell viability and invasiveness behaviors of B-CPAP cells. The first finding of our present study was the synergistic effects of CX and VPA on inducing apoptosis and decreasing viability of B-CPAP cells. The second finding was the synergistic effects of CX and VPA on inhibition of invasion and migration of B-CPAP cells in Transwell assay. Finally, we found that CX and VPA reduced the expression of MMP-2 at transcriptional level while had no significant effects on the expression of MMP-9.
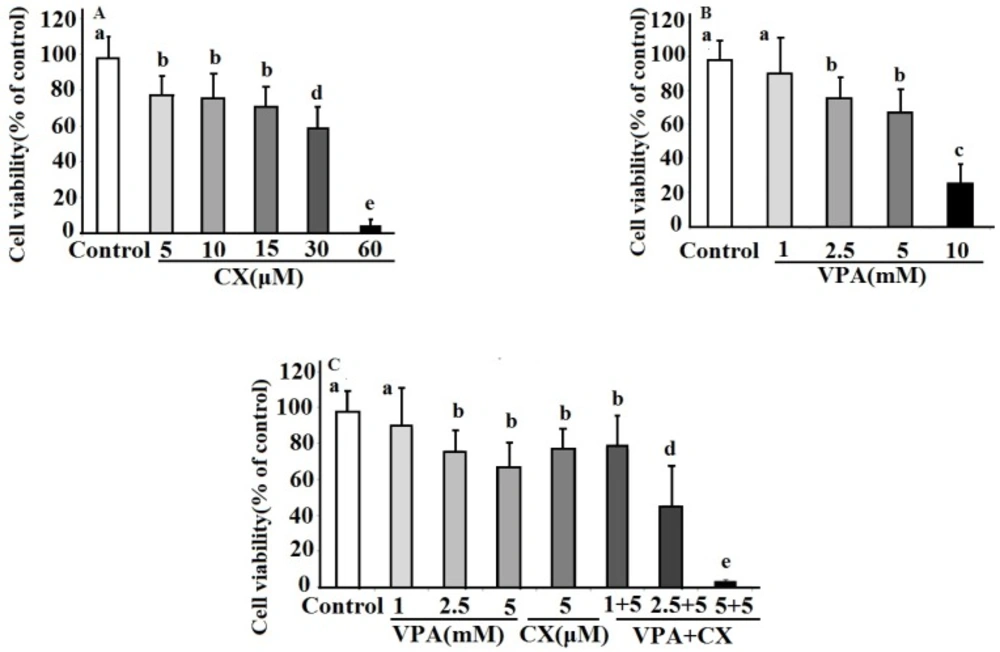
Findings of MTT assay in this study revealed a decrease in viability of B-CPAP cells following treatment with VPA in dose and time dependent manner which consistent with the findings of previous studies conducted on other tumor cell lines (
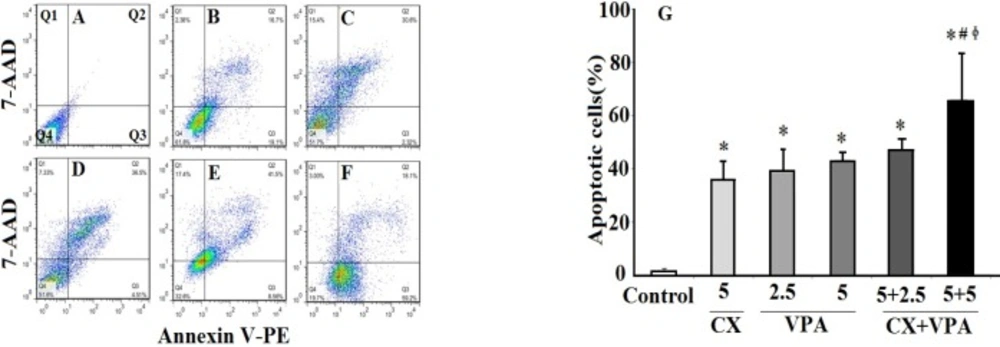
20). The lowest effective dose of VPA was 2.5 mM after treatment for 48 h. Flowcytometric analyses showed that VPA at 2.5 mM concentration induced apoptosis of B-CPAP cells, considerably. These results indicates that apoptosis is the main causes of VPA-induced cell death. In supporting our data, apoptotic effects of other HDACi such as trichostatin A and vorinostat have been demonstrated in previous studies (
21). Although the exact molecular mechanisms responsible for the apoptotic effects of HDACi remain elusive yet, up regulation of miR-129-5p expression (
12), increase in reactive oxygen species production (
22), inhibition of cell survival signaling pathways such as RAS/RAF/ERK and PI3K/AKT /mTOR (
23), and causing DNA double strand break DNA are among proposed mechanisms (
23).
The effects celecoxib (CX), sodium valproate (VPA) and combination of both drugs (VPA+CX) on viability of BCPAP cells using MTT assay. The represented data are mean ± SD of at least three independent experiments and were analyzed using one-way ANOVA followed by Tukey’s post-hoc test. In each figure, groups indicated with different letters (a, b, c, d and e) had statistical significant differences at P < 0.05.
The effects of VPA and CX on apoptosis of B-CPAP cells. Represented flowcytometric charts (A-F) show the percentage of live (Q4), early apoptotic (Q3), late apoptotic (Q2), and necrotic cells (Q1) in different group (A: control group, B: 5 µM of CX, C: 2.5 mM of VPA, D: 5 mM of VPA , E: 5 µM of CX + 2.5 mM of VPA, F: 5 µM of CX + 5 mM of VPA). Histogram (G) compares the percentage of total apoptotic cells (Q2+Q3) in CX, VPA, and CX+VPA -treated and control B-CPAP cells. The represented data are mean ± SD of at least three independent experiments. Data were analyzed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05 compared to control group; #P < 0.05 compared to CX (5 µM) group, ɸP < 0.05 compared to VPA (2.5 and 5 mM)-treated groups.
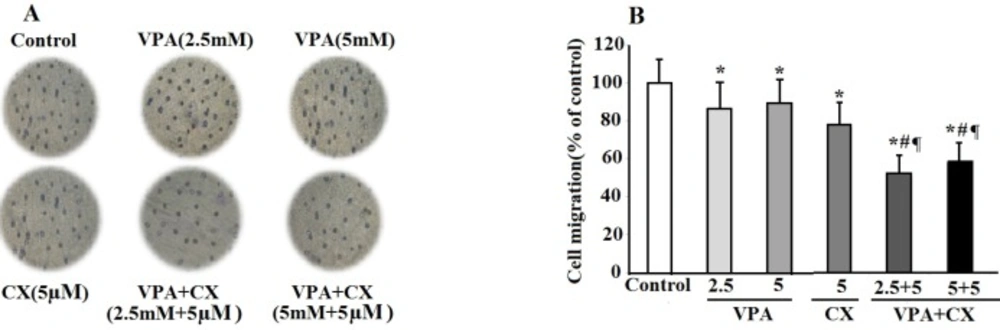
The effects of VPA and CX on the migration of B-CPAP cells in-vitro. (A) Representing light microscopy photomicrographs (40x) showing the migration ability of BCPAP cells in the control, VPA and CX-treated groups. (B) Histogram compares the mean ± SD of migrated cells in different group. Data was obtained from at least three independent experiments and were analyzed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05 compared to control group; #P < 0.01 compared to CX (5 µM); ¶P < 0.001 compared to VPA (2.5 and 5 mM)-treated cells.
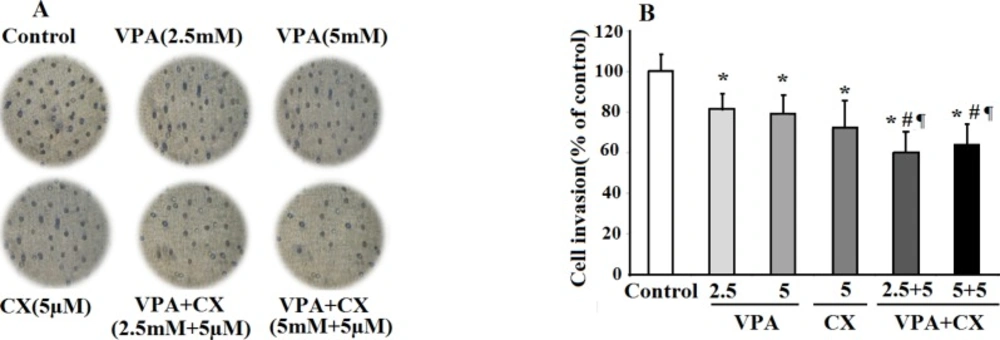
The effects of VPA and CX on the invasion of B-CPAP cells in-vitro. (A) Representing light microscopy photomicrographs (40x) showing the invasion ability of BCPAP cells in the control, drugs-treated groups. (B) Histogram compares the mean ± SD of invaded cells in different group. Data was obtained from at least three independent experiments and were analyzed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05 compared to control group; #P < 0.05 compared to CX (5 µM); ¶P < 0.01 compared to VPA (2.5 and 5 mM)-treated cells
(A) The effects of VPA and CX on the exoression of MMP-2 and (B) MMP-9 in B-CPAP cells. Gene expression was normalized to the level of GAPDH within each sample using the relative ΔΔCT method. The represented data are mean ± SD of at least three independent experiments and were analyzed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05 compared to control group.
Anti-cancer effects of selective COX-2 inhibitors including CX against numerous tumors have also been reported (
24). Our data in the present study showed that CX reduced B-CPAP cell viability in the MTT assay dose dependently, with the lowest effective dose of 5 µM. At this concentration, CX increased cell apoptosis to about 25 folds of non-treated cells. Results also revealed a significant inhibition of B-CPAP cells migration and invasion at 5 µM concentration of CX. A significant down regulation of MMP-2 was also observed at this concentration. These findings are consistent with the results of previous reports which revealed an increased expression of COX-2 and its association with increased invasiveness potential of PTC tumor cells (
25-
26) and also with the effects of another COX-2 inhibitors, NS-398, on suppression of PTC cells migration and invasion (
27).
Despite the beneficial effects of CX in the treatment of cancer, the major problem in the use of this drug is its side effects on cardiovascular system (
28). Thus, the use of CX in low concentrations and concurrent with other anti-cancer agents in combination therapy procedures has been suggested. In this context beneficial effects of CX in improving therapeutic efficacy of chemotherapeutic agents (
29); tyrosine kinase inhibitors (
30), radiotherapy (
31), and gene therapy (
32) have been demonstrated. Combination therapy using HDACi has also considered as an attractive therapeutic strategy for the treatment of cancer in recent years (
33). The synergistic effects of HDACi with a variety of therapeutic modalities including radiotherapy (
34) and chemotherapy (
35) have been demonstrated previously. Cha
et al. demonstrated synergistic effects of VPA and tumor necrosis factor-related apoptosis-inducing ligand in inducing apoptotic cell death of PTC cells (
13). In addition the synergy between HDACi and inhibition of NF-κB on proliferation of B-CPAP cells has also been demonstrated (
36).
In the present study, we conducted a combination therapy study using low concentrations of VPA and CX on cell survival and invasiveness properties of B-CPAP cell. Findings showed synergistic effects of this combination on cell death and apoptosis of B-CPAP cells. Furthermore, concurrent use of VPA and CX, decreased cell migration and invasion of B-CPAP cells compared to use of each drug alone. These findings are consistent with the observed synergistic effects of VPA and CX combination therapy against cell viability of neuroblastoma cells (
37). The exact molecular mechanisms that are behind the synergistic effects of VPA and CX in B-CPAP cells viability remain unclear. It has been reported that VPA and other HDACi suppressed the IL-1beta- induced COX-2 gene expression and protein expression in the endometriosis (
38). Thus, the observed synergistic effect between VPA and CX in our study might be due to their combined influences on COX-2 activity. In addition, it has been reported that both VPA and CX induced apoptosis of neuroblastome cells via up-regulation of Bax apoptotic protein and increase in Bax/Bcl2 ratio (
37). Further studies are required to explore the role such mechanisms in the synergistic effects of VPA and CX in B-CPAP cells.
While both of the drugs decreased the expression of MMP-2 alone, the result did not reveal any synergistic interaction between CX and VPA against MMP-2 expression when used in combination. Although the definitive explanation for this observation is not yet available, the stimulatory effects of VPA on a signaling pathway by which CX reduces MMP-2 expression may be a possible description for this finding. Wnt/β-catenin signaling pathway may be a good candidate in this issue. In this signaling pathway, free cytoplasmic β-catenin translocates into the nucleus and activates transcription of MMP-2 gene (
39). It has been demonstrated that CX reduced MMP-2 expression through inhibition of β-catenin accumulation in the nucleus (
40). In contrary to CX, VPA activates Wnt/β-catenin signaling pathway (
41). Taken together, it may hypothesize that VPA has two activities, on one hand it reduced MMP-2 expression and on the other hand it antagonize CX effect on MMP-2 expression. Further studies are necessary to elucidate the details of CX and VPA mechanisms in this issue.
In the present study, annexin V/7-AAD detection flow cytometric technique was applied as a reliable technique for accurate quantifying cell apoptosis. However, the mechanism of apoptotic effects of VPA and CX did not investigate, therefore, additional techniques for apoptosis assay such as analyses of caspases activation, determination of Bax/Bcl2 ratio, and DNA fragmentation assay are required to determine the mechanism underlying apoptotic effects of VPA and CX(
42). Moreover, the results of real time PCR showed a decreased level of MMP-2 upon drugs treatment, suggesting that VPA and CX probably affects B-CPAP invasiveness through inhibition of MMP-2. Further studies employing gelatin zymoraphy or direct measuring of MMPs activity and assay for MMPs inhibitor (
43) are suggested to explore the exact effects of VPA and CX on MMPs activity.
In conclusion our findings in this study suggest that CX and VPA could act synergistically at low concentration to increase B-CPAP cell death through induction of apoptosis. In addition, they have ability to reduce cell migration and invasion through inhibition of MMP-2 expression.