Botanical material
The leaves of Nymphaea lotus were collected from Ile Ife, Osun State, and Southwest Nigeria. They were authenticated by comparing with corresponding herbarium specimen (voucher number: LSH. 2012/6) deposited at the Lagos State University herbarium. The leaves were air dried and ground to powder prior to extraction.
Preparation of crude extract
Five kilograms of powdered plant material was macerated with 80% ethanol at room temperature. This was filtered and concentrated to dryness in vacuo at room temperature. The ethanolic extract (380 g) was suspended in water and successively partioned to give hexane extract (56g), dichloromethane (5.5 g), ethyl acetate (5g), butanol (7.4 g) and aqueous (131 g) fractions after removal of solvents in vacuo. The resulting fractions were stored at -10 °C until further use.
Drugs and Treatment
Kynuramine, Clorgyline, Deprenyl (Sigma-Aldrich, St. Louis, MO, USA), Tween 80 (2% Polyoxyethylenesorbitan monooleate), p-Chlorophenylalanine (PCPA) and Yohimbine (YOH) were purchased from Cristália, Itapira, SP, Brazil. Dimethyl Sulfoxide (DMSO; Sigma-Aldrich, St Louis, MO, USA), α-Methyl-p-tyrosine (AMPT; Sigma-Aldrich, St. Louis, MO, USA), Diazepam (DZP), Imipramine (IMI) and Prazosin (PRAZ) were purchased from Cristália, Itapira, SP, Brazil. Recombinant Human Monoamine Oxidase-A and –B (MAO-A and -B) were purchased from BD Biosciences Bedford, MA, USA. For in-vivo assay, drugs were dissolved in a vehicle [a mixture of 0.9% NaCl and 5% Tween-80 (v/v)] and administered (orally - p.o. or intraperitonealy - i.p) in a volume of 1 mL per 100 g of mice body weight. Drugs were dissolved in DMSO to yield a final DMSO concentration of 1.0% in the reaction mixture for in-vitro assay.
Pharmacological approaches
General pharmacological test
General pharmacological tests were conducted using a modified method of Malone [26] to detect general behavioral changes, effective doses and any signs of toxicity. Animals were treated through subcutaneous (s.c), intraperitoneal (i.p) or oral (p.o) route with CEN (5, 25, 125 or 625 mg/kg) or vehicle 10 mL/kg and observed periodically for 7 days.
Sodium pentobarbital sleep induction
Animals were treated orally with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or diazepam (1 mg/kg) one hour prior to intraperitoneal administration of sodium pentobarbital (50 mg/kg). Sleep latency (time to the loss of righting reflex) and duration (voluntary recovery of the righting reflex) were recorded.
Rota-rod
The effect on motor coordination was assessed using a rota-rod apparatus. The rota-rod apparatus consisted of a base platform and an iron rod of 3 cm diameter and 30 cm length, with a non-slip surface. Mice were selected randomly, subjected to oral treatment with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or diazepam (1 or 5 mg/kg) and placed individually to walk at 22rpm on the rotating rod one hour after treatment. Fall latency was recorded as an interval between the mounting of the animal on the rod and falling off of it.
Light/dark box test (LDB)
Mice were treated orally with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or diazepam (DZP 1 or 5 mg/kg) prior to assessment in LDB. The animals were placed at the center of the light area facing the opening of the dark area one hour after treatment. The number of transitions between the two compartments and the time spent in the light area were recorded for five min.
Elevated plus maze test (EPM)
Mice were treated orally with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or diazepam (DZP, 1 or 5 mg/kg) prior to assessment by elevated plus maze. One hour after treatment the animals were placed individually at the center of the plus maze and observed for five minutes. The number of entries into the open arms and time spent were recorded for statistical analysis.
Forced Swimming Test
Mice were treated orally with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or imipramine (IMI, 15 mg/kg) and subjected to swimming for six minutes (
27). The swimming duration and immobility periods were recorded. The immobility period was considered any time spent by the mouse floating and making passive movements necessary to keep its head afloat.
| Nymphaea spp. | Natural Product | Reference |
|---|
| N. alba | cyanidin 3-(6”-acetylgalactoside) (Figure.1 Gi); delphinidin 3-(2”-galloyl-6”-acetylgalactoside) (Figure.1 Gii); delphinidin 3-(6”-acetylgalactoside) (Figure.1 Giii); cyanidin 3-(2”-galloyl-6”-acetylgalactoside) (Figure.1 Giv); delphinidin 3-(2”-galloylgalactoside) (Figure.1 Gv); delphinidin 3-galactoside (Figure.1 Gvi); cyanidin 3-galactoside (Figure.1 Gvii) | (19) |
| N. ampla | 7,3’4’-trihydroxy-5-O-β-D-(2”-acetyl)-xylopyranosylisoflavone (Figure 1 Ai); 7,3’,4’-trihydroxy-5-O-α-L-rhamnopyranosylisoflavone (Figure 1 Aii); quercetin 3-rhamnoside (Figure.1 Bii); quercetin 3-xylopyranoside (Figure 1 Biii); quercetin 3-glucopyranoside (Figure 1 Biv); methyl gallate (Figure.1 C) | (23) |
| N. elegans | quercetin 3-rhamnoside (Figure 1 Bii); β-sitosterol (Figure 1 Fi); β-sitosteryl-3-O-β-D-glucopyranoside (Figure 1 Fii) | (23) |
| N. gracilis | methyl 3-O-β-D-glucopyranosyloleanolate, (E) 28-O-β-D-glucopyranosyloleanolate (Figure 1 Dii); 28-O-β-D-glucopyranosyl-oleanolate (Figure 1 E); β-sitosterol (Figure 1 Fi); β- sitosteryl -3-O-β-D-glucopyranoside (Figure 1 Fii). | (23) |
| N. lotus | myricitrin (Figure 1 I); 1,2,3,4,6-pentagalloyl-D-glucose (Figure 1 L); nympholide A (Figure 1 J); nympholide B (Figure 1 K);myricetin-3’-O-(6”-p-coumaroyl)glucoside (Figure 1 M). | (16,21) |
| N. pulchella | 7,3’4’-trihydroxy-5-O-β-D-(2”-acetyl)-xylopyranosylisoflavone (Figure 1 Ai); 7,3’,4’-trihydroxy-5-O-α-L-rhamnopyranosylisoflavone (Figure 1 Aii); kaempferol 3-rhamnopyranoside (Figure 1 Bi); β-sitosterol (Figure 1 Fi); β- sitosteryl-3-O-β-D-glucopyranoside (Figure 1 Fii) | (23) |
| N. x marliacea | myricetin 3-O-α-L-rhamnopyranosyl(1→6)β-D-galactopyranoside (Figure 1 H); delphinidin 3-(2”-galloyl-6”-acetylgalactoside) (Figure 1 Gii); delphinidin 3-(6”-acetylgalactoside) (Figure 1 Giii); delphinidin 3-galactoside (Figure 1 Gvi) | (22) |
| N. x marliacea var. Escarboucle | delphinidin 3-(2”-galloyl-6”-acetylgalactoside) (Figure 1 Gii); delphinidin 3-(6”-acetylgalactoside) (Figure 1 Giii); cyanidin 3-(2”-galloyl-6”-acetylgalactoside) (Figure 1 Giv); delphinidin 3-(2”-galloylgalactoside) (Figure 1 Gv); delphinidin 3-galactoside (Figure 1 Gvi) | (22) |
| Sample | Unit | MAO-A IC50 | MAO-B IC50 |
|---|
| CEN | μg/mL | 149.503 ± 0.05 | - |
| Clorgyline | μM | 0.024 ± 0.03 | - |
| Deprenyl | μM | - | 0.072 ± 0.08 |
| Treatment (p.o) | [BDNF] pg/mg total protein |
|---|
| Vehicle 10 mL/kg | 80.32 ± 3.50 |
| CEN 20 mg/kg | 70.72 ± 5.250 |
| CEN 60 mg/kg | 84.33 ± 2.20 |
| CEN 180 mg/kg | 86.05 ± 6.10 |
Natural products isolated from Nymphaea spp. (A) 5-glycosyl isoflavones, (B) 3-glycosyl flavones, (C) methyl gallate (D) methyl 3-O-β-D-glucopyranosyloleanolate, (E) 28-O-β-D-glucopyranosyloleanolate, (F) sitosterols, (G) 3-O-glycosylated anthocyanins, (H) myricetin 3-O-α-L-rhamnopyranosyl(1→6)β-D-galactopyranoside, (I) myricitrin (J) nympholide A, (K) nympholide B, (L) 1, 2, 3, 4, 6-pentagalloyl-D-glucose, (M) myricetin-3’-O-(6”-p-coumaroyl) glucoside
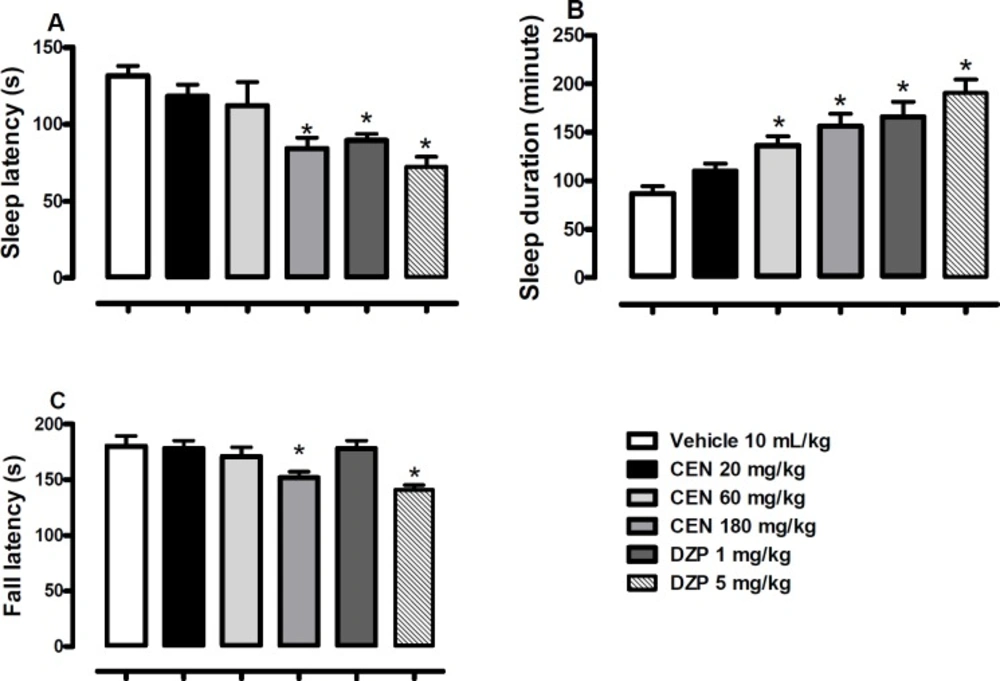
Effect of vehicle, crude leaf extract of Nymphaea lotus (CEN) 125, 250 or 500 mg/kg or diazepam (DZP) 1 or 5 mg/kg or on latency (A), duration (B) of sodium pentobarbital (60 mg/kg) induced hypnosis, and latency of fall on the Rota-rod (C). Results are expressed as mean ± SEM; n = 10 in each group. * indicate p < 0.05 as compared with vehicle treated group (One-way ANOVA followed by Dunnett´s test
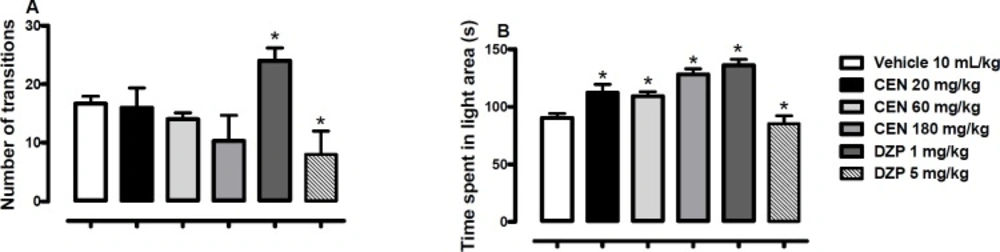
Effect of oral administration of vehicle 10 mL/kg, crude leaf extract of Nymphaea lotus (CEN) 20, 60 or 180 mg/kg, diazepam (DZP) 1 or 5 mg/kg on the number of transitions (A) and time spent in the light area of light/dark box (B). Results are expressed as mean ± SEM; n = 10 in each group; *p < 0.05 vs vehicle treated group (One-way ANOVA followed by Dunnett´s test
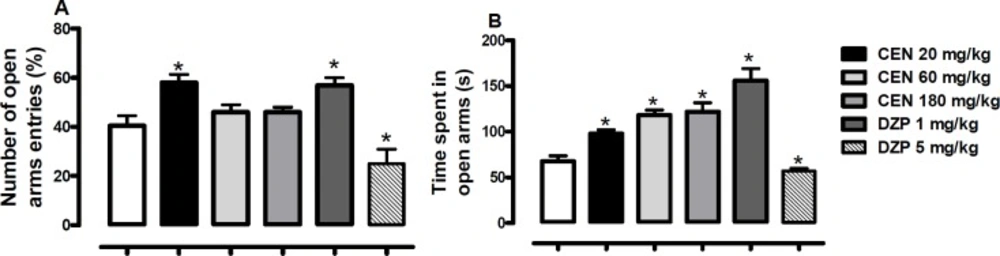
Effect of oral administration of vehicle 10 mL/kg, crude leaf extract of Nymphaea lotus (CEN) 20, 60 or 180 mg/kg, diazepam (DZP) 1 or 5 mg/kg on the (A) percentage of entries and (B) time spent in the open arms of the elevated plus maze. Results are expressed as mean ± SEM; n = 10 in each group; *p < 0.05 vs vehicle treated group (One-way ANOVA followed by Dunnett´s test
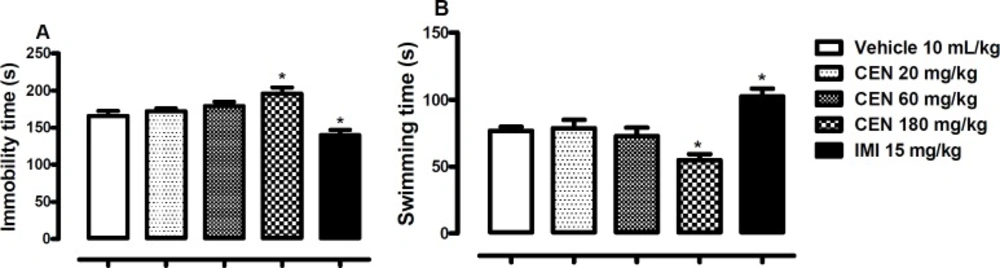
The effect of oral administration of vehicle 10 mL/kg, crude leaf extract of Nymphaea lotus (CEN) 20, 60 or 180 mg/kg, or imipramine (IMI) 15 mg/kg on the immobility (A) and swimming time in the FST (B). Each column represents the mean ± SEM of 10 animals. *p < 0.05 vs vehicle treated group. (One-way ANOVA followed by Dunnett´s test
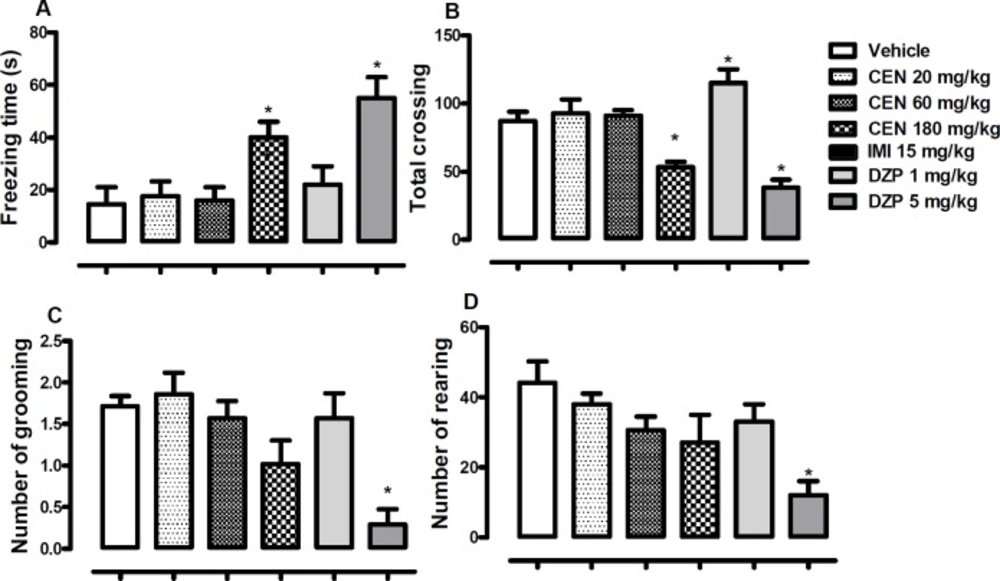
The effect of oral administration of vehicle 10 mL/kg, crude leaf extract of Nymphaea lotus (CEN) 20, 60 or 180 mg/kg, or imipramine (IMI) 15 mg/kg on the (A) freezing time, (B) total crossing, (C) number of grooming and (D) number of rearing in the open field test. Each column represents the mean ± SEM of 10 animals. *p < 0.05 vs vehicle treated group. (One-way ANOVA followed by Dunnett´s test
Showed effect of CEN 20 mg/kg or vehicle 10 mL/kg (p.o) after pretreatment
Open field exploratory activity
Mice were treated orally with vehicle (10 mL/kg), CEN (20, 60 or 180 mg/kg) or diazepam (DZP, 1 or 5 mg/kg) and then placed into a circular open field (50 cm high walled area whose 62.80 cm
2 floor was divided into eight equal sectors). Parameters of total crossing, freezing time, number of grooming and rearing activities were scored during five minute test periods (
27). The apparatus was cleaned with 10% alcohol at the end of each test.
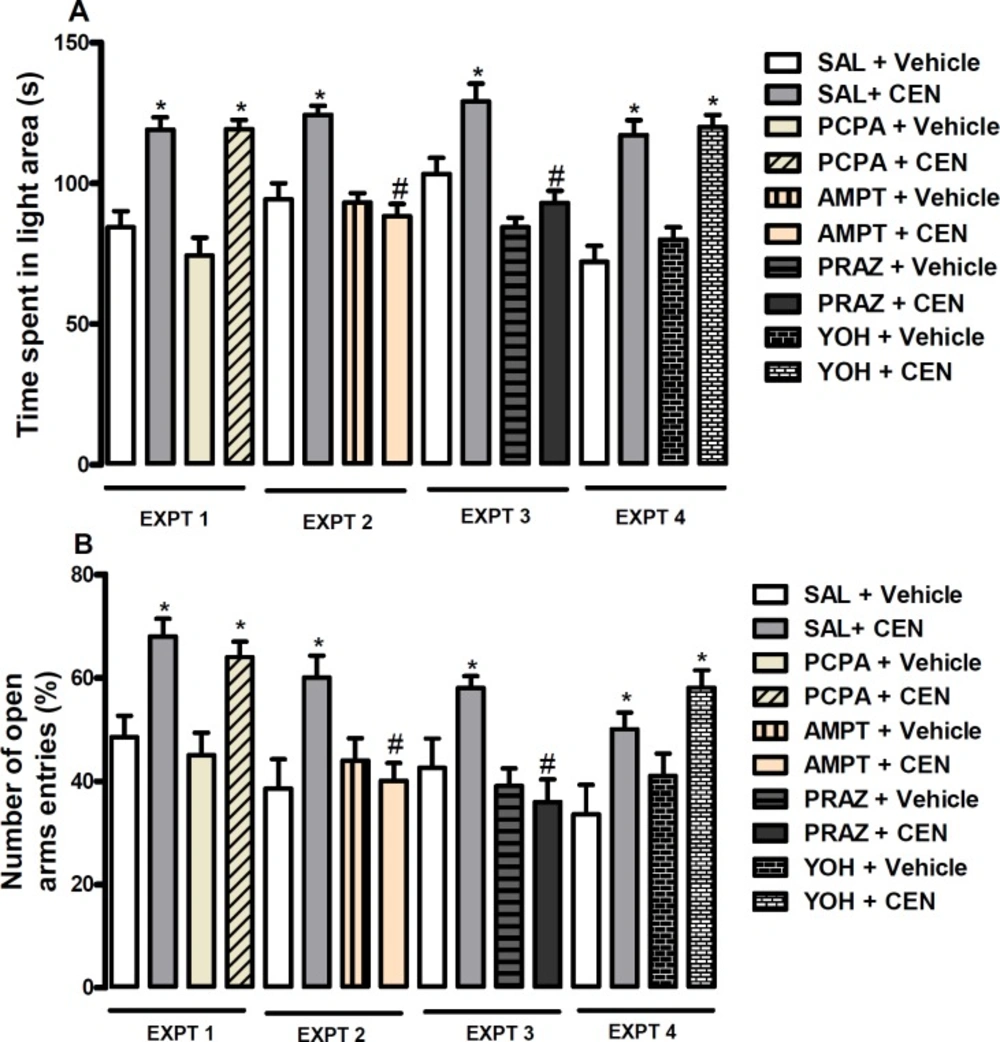
Mechanism of anxiolytic like effect of CEN
Mice were pretreated intraperitoneally with NaCl 0.9% - SAL or PCPA 100 mg/kg (experiment-EXPT 1), NaCl 0.9% - SAL or AMPT 100 mg/kg (experiment-EXPT 2), NaCl 0.9% - SAL or PRAZ 0.5 mg/kg (experiment-EXPT 3) or NaCl 0.9% - SAL or YOH 1 mg/kg (experiment-EXPT 4) prior to oral treatment with vehicle or CEN 20 mg/kg. One hour post-treatment animal were evaluated using a forced swimming test.
Inhibition assay using recombinant human MAO-A and -B
A monoamine oxidase (MAO) inhibition assay utilizing kynuramine as the substrate was employed to investigate the potential activity of CEN on recombinant human MAO-A and MAO-B (
28). CEN was assessed for inhibitory activity at a test range of 1-100 mg/mL, with clorgyline (10
-10 - 10
-5 M) and deprenyl (10
-10 - 10
-5 M) being employed as MAO-A and MAO-B selective inhibitors, respectively. The substrate concentration of 80 and 50 μM kynuramine was chosen for MAO-A and -B, respectively, based on the previous substrate concentration at half V
max (
29). Inhibition was calculated as percent of product formation compared to the corresponding control (enzyme-substrate reaction) without the inhibitors. The reactions were carried out in 0.1 M potassium phosphate buffer at pH 7.4, with incubation mixtures containing 5µg/mL of MAO-A or 12.5 µg/mL of MAO-B. The total reaction volume was 200 µL yielding a final DMSO concentration (vehicle) of 1.0% in the reaction mixture. Reaction mixtures were pre-incubated for 10 minutes at 37 °C prior to the addition of MAO-A or MAO-B to measure enzymatic activity. The reactions were stopped by the addition of 75 µL of 2N NaOH. The formation of 4-hydroxyquinoline was determined fluorometrically using a SpectraMax M5 fluorescence plate reader (Molecular Devices, Sunnyvale, CA) with 320 nm excitation and 380 nm emission wavelength calculated by the Soft Max Pro program.
Hippocampal protein extraction and BDNF quantification
Mice were treated orally of or vehicle 10 mL/kg, CEN 20, 60 or 180 mg/kg and sacrificed (after one hour) to collect their hippocampus. Hippocampal tissues were homogenized in a lysis buffer (NaCl 1mM; EDTA 4 mM, Tris-HCl 100 mM; albumin 2%, Triton-X 100 2%, thrimerosal 0.01%, pH 7.0, glycerol 10%, protease inhibitor cocktail - GE) in a 1:40 w/v ratio. After centrifugation (16800 x g, 4 °C, 35 min.), the supernatant was collected and stored at -80 °C. Hippocampal brain derived neurotrophic factor (BDNF) was measured using an ELISA kit (BDNF Emax® ImmunoAssay System kit, Promega, Madison, WI, USA) according to the manufacturer’s instructions.
The total protein level of sample was normalized and measured using the Bradford method
Statistical analysis
Parametric data are expressed as means ± SEM following appropriate statistical analysis, specifically: unpaired Student’s
t-test, one way ANOVA followed by the Dunnett´s test as post hoc test or two way ANOVA followed by the Bonferroni as post hoc test. Significance difference between or among groups were set at
p < 0.05 (
31). GraphPad Prism statistical software version 6.03 for Windows (GraphPad Software, Inc., San Diego, CA, USA) was used for data analysis.