Chemicals
Diazinon (DZN) from Merck Co.(Germany) (99% purity), pregnant mare’s serum gonadotrophin (PMSG) from Intervet Inc.(Germany), human chorionic gonadotropin (hCG) from Intervet Inc.(Germany), RNX plus solution from Sinaclon (Iran), cDNA first strand synthesis kit from Fermentase (Germany), and SYBR Green Premix 2X from Takara (Japan) were used in this study.
Animals
Immature Wistar female rats were obtained from the Pastor Institute (Experimental Animal Center, Shiraz, Iran) and held in our laboratory. The animals were fed a standard laboratory diet and water ad libitum and housed in a temperature-controlled room (22 ± 2 °C) in cages with a 12 h light-dark cycle, relative humidity of 50–55%, then all studies were begun when the animals were 24 days old (36-38 g). All procedures used were approved by the University Animal Care and Use Committee, Shiraz University of Medical Sciences. The procedures were performed in accordance with institution guidelines for animal care and use.
Experimental design
Fifty Immature 24-day-old Wistar rats were randomly divided into 5 time groups; four time groups comprised of 10 rats, 5 as control and 5 as DZN treatment for each time point. One time group was considered without injection before ovulation time (8 h post-hCG). All the animals employed in this study received intra-peritoneal (I.P) injection with PMSG (15 IU) to stimulate follicular maturation. Forty-eight h later, the rats were injected with hCG (15 IU, I.P) to induce ovulation process. DZN was dissolved in corn oil to consistent absorption and administered in a single dose (70 mg/kg body weight, I.P); control animals received only the vehicle under the same condition; at 12 h post-hCG injection (around ovulation time). The LD50 of DZN in female rats is 300 mg/kg body weight (
38), which was taken as the reference value. The reason for selecting a dose of 70 mg/kg bw in the present experiment is selected based on pilot study in our laboratory to determine its sub lethal dose that caused toxicity to the animals and simultaneously did not cause mortality of the animals.
The rats were sacrificed by spinal dislocation at defined time points at 4-h intervals from 8 to 24 h post-hCG injection. The ovaries were rapidly removed, washed in a cold saline solution, snap-frozen in liquid nitrogen and then stored at -80 °C for RNA extraction.
Histological analysis
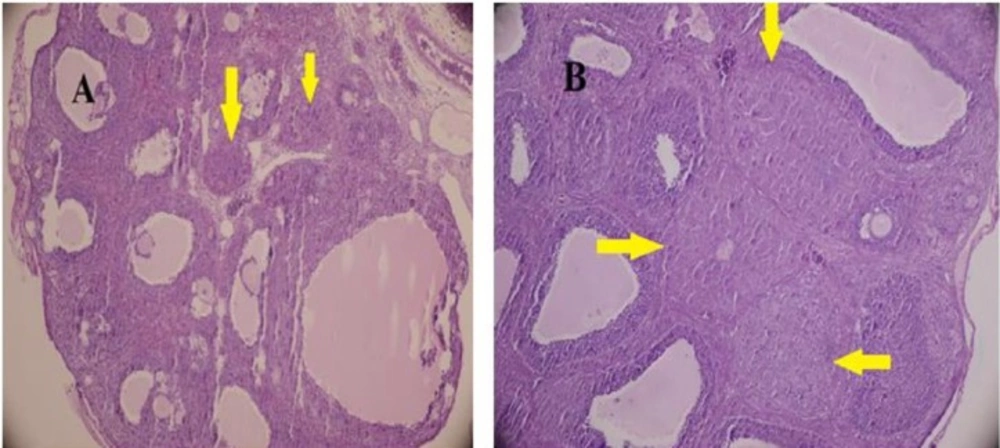
The ovaries were dissected and fixed in Bouin’s fixative for histological examination. The fixed tissue was cut into 5-7-μm thick sections and stained with hematoxylin and eosin.
For each ovary, at least six independent sections were selected, and the total number of CL and Graafian follicles was counted. At least 100 CL and follicles (FL) in different stages were counted and the fraction of FL/CL was calculated for each case.
The follicles were classified as Secondary when they had more than two layers of granulosa cells and labeled as antral when the follicle contained fluid. In the Graafian follicle, the oocyte occupied an excentric position and the antral cavity was filled with fluid.
The size of CL was measured by computer-assisted morphometric program (Olysia, Olumpus).
Preparation of total RNA and cDNA syntetase
Total RNA was extracted from approximately 100 mg of ovary tissue by RNX plus solution (Sinaclon, Iran) in a clean RNase-free tube. Concentration and purity of RNA were quantified by NanoDrop ND-100 spectrophotometer (Thermo Scientific, Waltham, MA, USA) at 260 nm and 280 nm.
RNA was converted to cDNA after treating with DNase I. Reverse transcription of RNA was done in a final volume of 20 μL by using cDNA first strand synthesis kit (Fermentase, Germany) by oligo (dT) primer
Quantitative Real-Time PCR
The sequence of the gene was obtained from Gen Bank and the primers were used in previous studies (
39,
40). The sequences of the primers are shown in
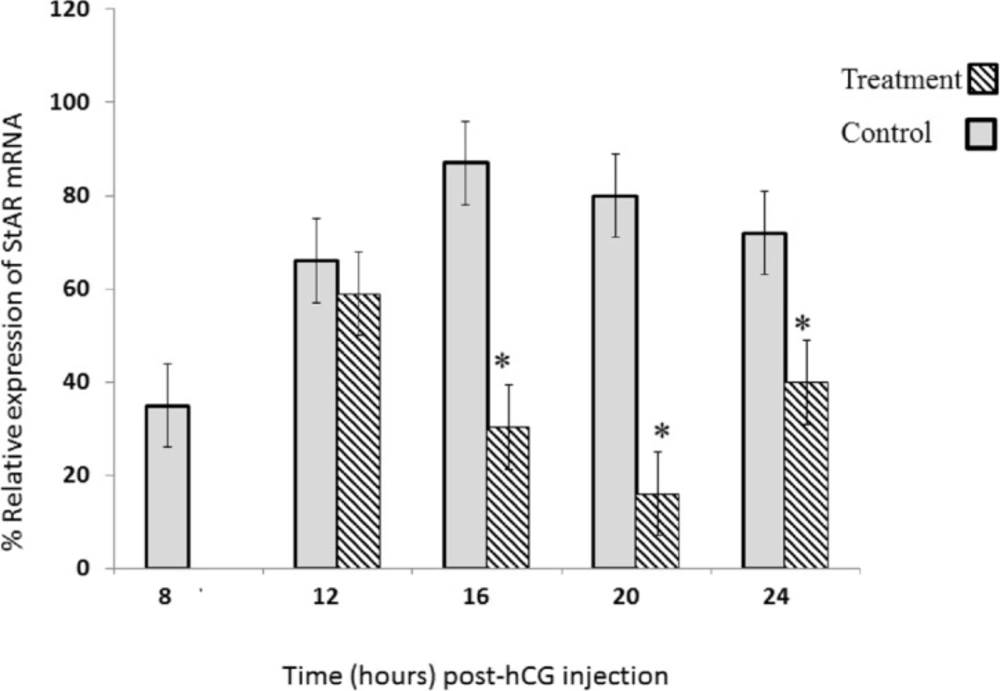
Table 1. For real-time quantitative PCR, 500 ng of RT product was used in whole volume of 25 μL containing 7.5 μL of SYBR Green Premix 2X (Takara, Shiga, Japan) and 10 PM of mix primer. Thermocycling conditions were; heat hold at 94 °C for 1 min followed by 40 cycles of denaturation at 94 °C for 20 sec, annealing and extention at 61 °C for 30 sec. We used the ΔΔ
CT method for determination of relative StAR gene expression. The
Ct of sample was compared with the
Ct of its internal control (β-actin). Real-time PCR was done with a BioradBiosystemsIQ5 detection system. All reactions were done in duplicate. Specificity of PCR reaction was double-confirmed by electrophoresis and melting curve analysis.
Statistics
Statistical analysis of data was carried out using SPSS (version 18) software. The results are expressed as mean ± SE of five experiments. One-way analysis of variance (ANOVA) and Bonferroni multiple comparison tests were used to test the difference between groups. P < 0.05 for comparison between study groups was taken as statistically significant.