All animal study protocols were approved by the animal care and use committee of the Kerman Medical University (Ethics Code: KNRC/92/6). Male Wistar rats (8-10 weeks old, 180-220 g) were housed under standard conditions (12/12 light dark cycle) throughout the study.
Experimental groups
In the present study, 80 male Wistar rats were randomly allocated into 8 experimental groups. The rats subjected to BRI/reperfusion were divided into four groups of n = 10, in which 0.5 mL of normal saline alone or normal saline containing EPO (1000 U/kg) was i.p. injected 30 min before the induction of renal ischemia followed by the reperfusion periods of 24 h (BRI-24 h +V group and BRI-24 h +EPO group, respectively) and 1 week (BRI-1w+V group and BRI-1w +EPO group, respectively). Sham-operated rats were also divided into four groups of n = 10 that received 0.5 mL of normal saline alone or normal saline containing EPO (1000 IU/kg) with periods equivalent to reperfusion of 24 h (sham-24 h+ V group and sham-24 h+ EPO) group, respectively) or 1 week (sham-1w +V group and sham-1w +EPO group, respectively).
Surgery and experimental protocol
Each rat was anesthetized deeply by ether and the abdominal region was shaved, soaked with betadine and covered with a sterile draper, leaving only its abdomen exposed for a midline electrosurgical laparotomy. In rats subjected to renal ischemia/reperfusion, there was occlusion of the right and left renal arteries and veins for 1 h in the BRI +V group by using a non-traumatic clamp, occlusion was verified visually by a color change of the kidney to a paler shade, and reperfusion was verified by a darkening.
In the sham +V group, all surgical procedures were performed but renal pedicles were only manipulated, and were maintained under anesthesia for the duration of the experiment. The abdominal incision was sutured at two layers by 2-0 silk. The whole surgical procedure was performed under sterile condition, and the rat was allowed to recover from the anesthesia prior returning to an individual cage. The surgical protocol was approved by the Shiraz University of Medical Sciences.
Treatment
The animals received saline (i.p.), or EPO (1,000 U/kg, single dose, (i.p) 30 min before surgery. EPO was provided as a gift from Pouyesh Darou product Company. The dose of EPO used was based on that found to be maximally effective in our previous published study (
26).
Assessment of Renal Function
At the end of the experiments, 1 mL blood samples were collected from anesthetized rats 24h and 1w after the reperfusion period. Plasma concentrations of BUN (blood urea nitrogen) and creatinine (Cr) were used as a marker of renal function. The measurements were performed by a lab technician blind to the groups.
Behavioral assessments
Passive Avoidance (PA) Learning
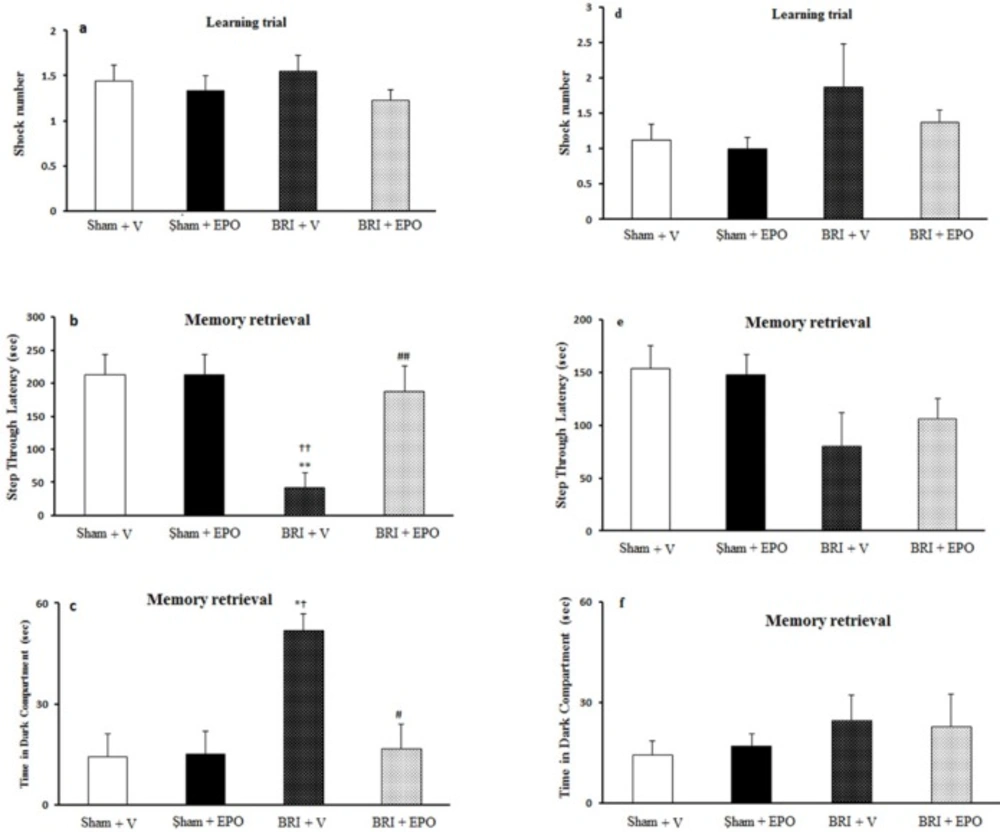
This test was used to evaluate fear learning in AKI rats and the possible effect of EPO on 24 h and 1w after reperfusion. The apparatus was a shuttle-box device with dimensions of 100*25*25 which consisted of two compartments, one dark and one illuminated part separated by a guillotine door, and a stainless rod grid serving as the base. The adaptation phase was followed by a single trial in which the animals were placed in the lit arena, after 10s, the door was opened and the animal was allowed to go to the dark compartment. Then the door was closed without electric shock and after 20s, the animal was placed in the cage. If the rats did not enter the dark chamber within 60s, they were eliminated from the test and replaced with a new rat. After 30 min of adaptation, for the learning trial this procedure was repeated. A mild shock was immediately administered (0.5 mA, 1.5 s) to the rat the third time that the animal entered the dark sector. This procedure was repeated until the animal did not enter the dark box in a 60s period. The number of shocks received was recorded in this phase.
In the retention trials, 24h after training, the test was performed to evaluate memory; in this step the animal was placed in the light arena and the latency to step into the dark sector as retention time (step through latency (STL), Time in dark compartment (TDC) and number of entrance into dark sector were measured as indicators of contextual memory) was recorded to a maximum of 300s (
27).
Morris Water Maze (MWM)
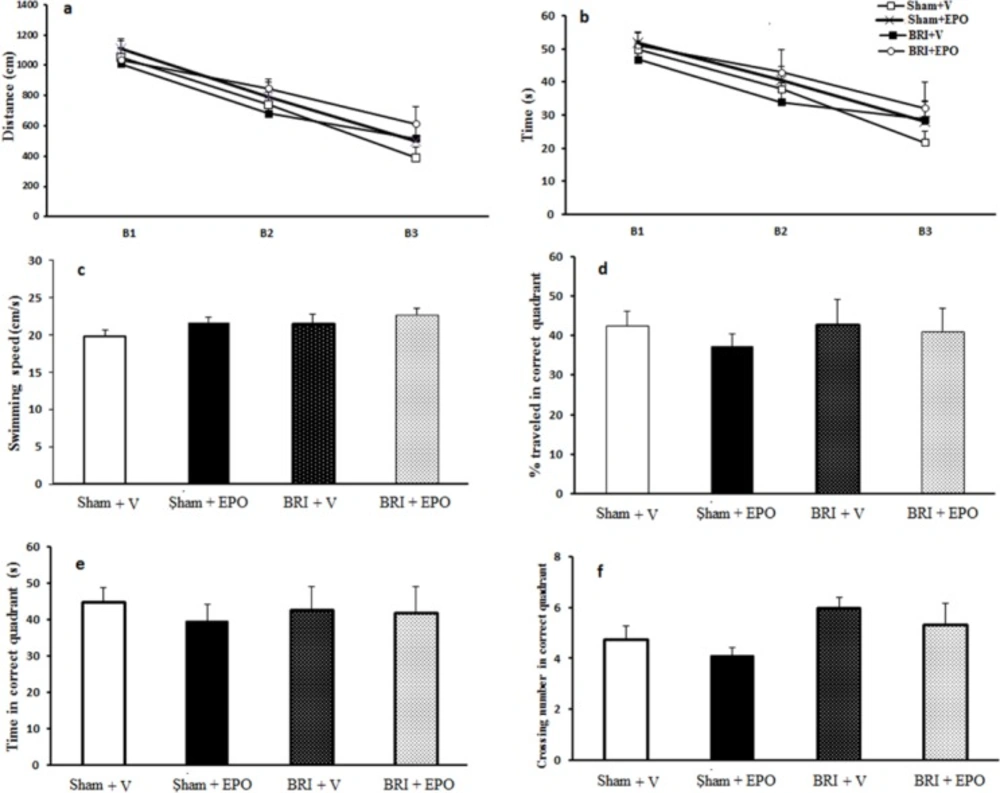
To assess spatial learning and memory, the animals were tested by a MWM as previously described (
28). Briefly, the test chamber was a circular tank (140 cm diameter, 45 cm height, a black pool), surrounded by extra-tank visual cues. A visible or submerged platform (15 cm wide, 35 cm height) was placed 1.5 cm above or below the water surface. Water temperature was maintained among 21–23 °C.
Rats’ behavior was recorded with the Ethovision system. The following parameters were recorded for each rat: total distance and time spent to reach the platform in three consecutive trials, number of crosses in the correct quadrant in the retention phase, percentage of time, and distance traveled in the correct quadrant.
In the training phase, each rat underwent three blocks of trials, each comprising four trials (inter-trial interval = 30 s). In each trial, animals were placed in one of the four quadrants facing towards the maze. Each rat was given 60s to find the platform, and if they did not find it, they were put on the platform by the examiner. After 30 s, the rat was again put to the trial.
After 2 h of the last block, the rats underwent a probe trial during which the platform was removed from the tank and the number of crosses in the correct quadrant and total time spent in target quadrant was recorded and analyzed for each rat.
Histological Assessment
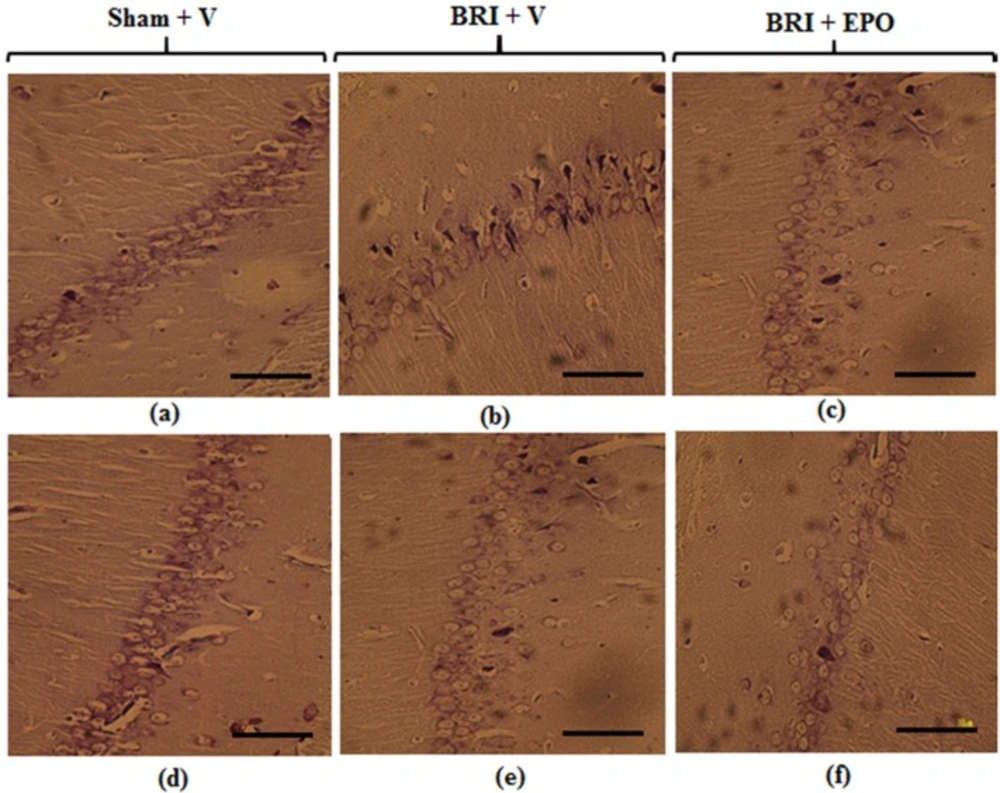
The rats were decapitated and their brains were removed 24 h and 1w after reperfusion. After tissue processing, paraffinzed brains were cut onto 6µm sections on a rotary microtome and the sections were stained with cresyl violet (Nissl staining) (
29).
Molecular experiment
Tissue dissection and preparation for Western blot
For molecular experiment, the rats were allocated in sham +V, BRI +V, and BRI +EPO groups. All of the animals were anesthetized with atmosphere CO
2 and decapitated 24 h after surgery. Their brains were rapidly extracted and placed on ice. The both whole hippocampi were freshly harvested and maintained in a microtube at −80 °C until homogenization for further western blot assay (
30).
BDNF immunoblot analysis
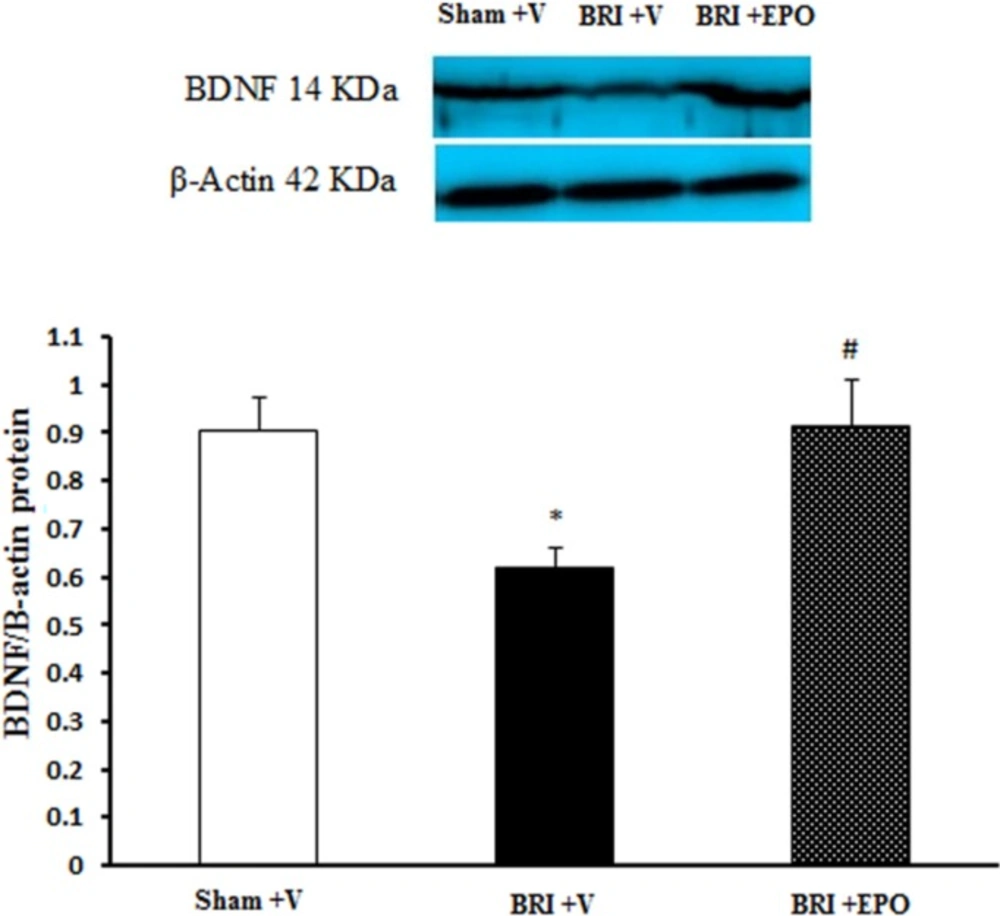
Dissociated hippocampus tissues were homogenized in ice-cold buffer containing 10 mM Tris–HCl (pH 7.4), 1 mM EDTA, 0.1% SDS, 0.1% Na-deoxycholate, 1% NP-40 with protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 2.5 μg/mL of leupeptin, 10 μg/mL of aprotinin) and 1 mM sodium orthovanadate (a phosphatase inhibitor). The homogenate was centrifuged at 15,000 rpm for 20 min at 4 °C. The resulting supernatant was retained as the whole cell fraction. Protein concentrations were measured using the Bradford method (Bio-Rad Laboratories, Muenchen, Germany). Equal amounts of protein were resolved electrophoretically on a 9% SDS polyacrylamide gel electrophoresis (SDS-PAGE) SDS-PAGE gel and transferred to PVDF (polyvinylidenefluoride) membranes. After blocking (overnight at 4 °C) with 5% non-fat dried milk in Tris-buffered saline with Tween 20 (blocking buffer, TBS-T, 150 mM NaCl, 20 mM Tris–HCl, pH 7.5, 0.1% Tween 20) for 2 h, at room temperature and then, the membranes were incubated overnight by a primary rabbit polyclonal antibody for BDNF (1:1000, sc-20981; Santa Cruz Biotechnology, Santa Cruz, USA) at 4 °C. After washing in TBS-T buffer (three times for 5 min each, at room temperature) the blots were incubated for 2 h at room temperature with an anti-rabbit IgG secondary antibody conjugated with horseradish peroxidase (1:15,000; GE Healthcare Bio-Sciences). Both primary and secondary antibodies were diluted in blocking buffer. The antibody-antigen complexes were revealed using the ECL system (Amersham Biosciences) and images were captured on a Gel Doc imaging system (Bio-Rad, Hercules, CA, USA), converted to a tiff file, and analyzed with Lab Work analyzing software (UVP, UK) was used to analyze the intensity of the expression. β-actin immunoblotting (antibody from Cell Signaling Technology, INC. Beverly, MA, USA; 1:1000) was used to control for loading (
30).
Statistical Analysis
Statistical procedures were performed using the SPSS software (version 18). ANOVA followed by Tukey’s post-hoc analysis was used to compare the differences between groups. The PA learning and probe data of MWM were analyzed by one-way ANOVA. Repeated measures ANOVA was used to analyze the data of MWM task in the learning phase.
The band density values were expressed as BDNF/β-actin ratio for each sample. The averages for different groups were compared using one-way ANOVA. All data were expressed as mean ± SEM. P <0.05 was considered statistically significant.