It is known that the classes and concentrations of plant chemical compounds vary in reason of the biotic and abiotic influences (
23), so initially the phytochemical analysis in order to verify if the extract contains bioactive chemical compounds similar described in other extracts.
Phytochemical analysis by Matos method revealed the presence of various compounds such as alkaloids, flavones, flavonols, xanthones, flavanonols, flavonones, and condensed tannins. Similar results were obtained by Bento
et al. (
10) that identified the presence of flavonoids, alkaloids, and tannins in the leaves of
Annona muricata, and Oviedo
et al. (
7) detected the presence of flavonoids and alkaloids. Other studies confirmed the presence of flavonoids and alkaloids in other species of the
Annona genus (
9).
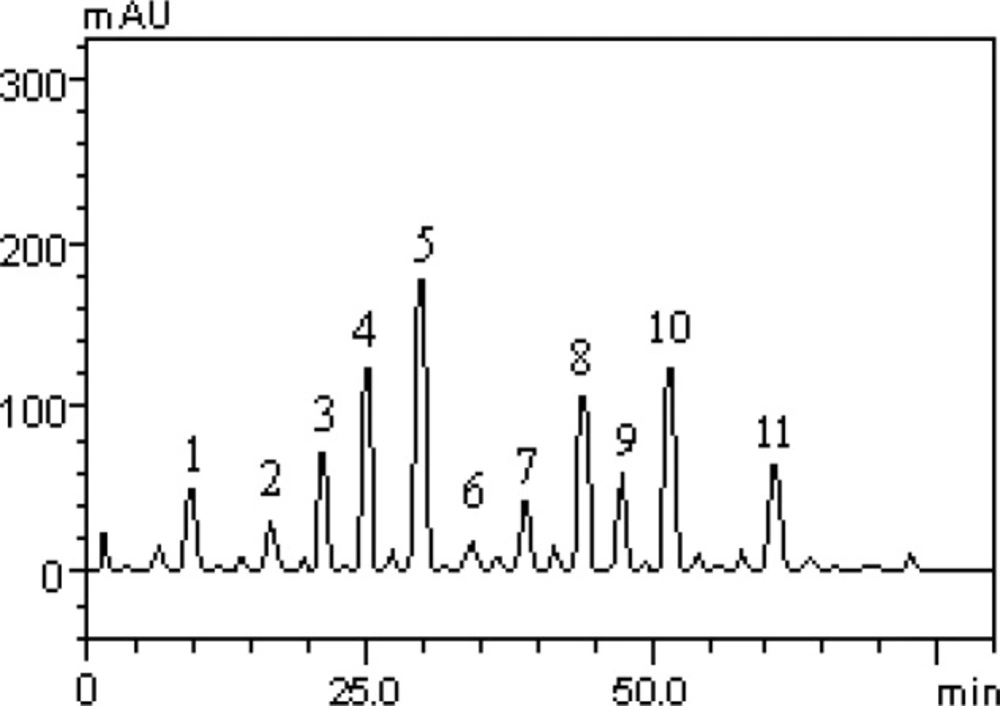
However, this is the first study to describe the quantification of flavonoids and phenolic acids of HLEAM by HPLC method. The results revealed the presence of the gallic acid (t
R = 10.09 min; peak 1), catechin (t
R = 16.32 min; peak 2); chlorogenic acid (t
R = 20.57 min; peak 3), caffeic acid (t
R = 24.93 min; peak 4), ellagic acid (t
R = 29.86 min; peak 5), epicatechin (t
R = 34.19 min; peak 6), rutin (t
R = 39.05 min; peak 7), isoquercitrin (t
R = 44.89 min; peak 8), quercitrin (t
R = 47.61 min; peak 9), quercetin (t
R = 51.07 min; peak 10) and kaempferol (t
R = 61.20 min; peak 11) (
Figure 1 and
Table 1).
Representative high performance liquid chromatography profile of HLEAM, detection UV was at 325nm. Gallic acid (peak 1), catechin (peak 2), chlorogenic acid (peak 3), caffeic acid (peak 4), ellagic acid (peak 5), epicatechin (peak 6), rutin (peak 7), isoquercitrin (peak 8), quercitrin (peak 9), quercetin (peak 10) and kaempferol (peak 11). Chromatographic conditions are described in the Methods section
| COMPOUNDS | HLEAM mg/g | LOD g/mL | LOQ g/mL |
|---|
| Gallic acid | 5.31 ± 0.02 a | 0.015 | 0.049 |
| Catechin | 2.79 ± 0.01 b | 0.032 | 0.105 |
| Chlorogenic acid | 6.25 ± 0.01 c | 0.009 | 0.029 |
| Caffeic acid | 10.14 ± 0.03 d | 0.024 | 0.078 |
| Ellagic acid | 13.07 ± 0.02 e | 0.013 | 0.042 |
| Epicatechin | 1.83 ± 0.01 f | | |
| Rutin | 5.20 ± 0.01 a | 0.027 | 0.090 |
| Isoquercitrin | 9.64 ± 0.03 g | 0.008 | 0.026 |
| Quercitrin | 6.19 ± 0.01 c | 0.035 | 0.114 |
| Quercetin | 10.21 ± 0.02 d | 0.019 | 0.063 |
| Kaempferol | 6.27 ± 0.03 c | 0.026 | 0.085 |
Some genera of Annonaceae produce weakly polar flavonoids which facilitate their diffusion of such compounds across the blood-brain barrier and so to act on the central nervous system (
24). Some studies indicate that flavonoids quercetin and kaempferol, identified in HLEAM, showed effects sedative (
25,
26) which may justify the use of this plant in the relief of anxiety, depression, and sleep disorders.
In the present study, a pharmacological screening and an acute toxicity test were assessed to evaluate the pharmacological and toxicological profile of HLEAM. The animals treated with HLEAM, i.p., at doses of 500, 1000, 1250, 1500, 1750, and 2000 mg/kg showed central nervous system depressant effect, such as sedation and decreased motility. The behavioral changes were evident 30 min after administration of the extract. At the dose of 1000 mg/kg, 50% of the animals died. With increasing doses of the extract (1250, 1500, 1750 mg/kg) the percentage of mortality increased to 75%. Finally, with the administration of an HLEAM dose of 2000 mg/kg, 100% of the treated animals came to death within 30 min after administration. The LD
50 was 1091.7 mg/kg, determined by the probit method. In folk medicine, several plants are used in the treatment of psychiatric disorders; this is due to the false belief that by being natural they are safer than synthetic psychotropic medications (
6).
However, the use of natural medication does not mean absence of side effects or toxicity; in this aspect, Caparros-Lefebvre
et al. (
27) showed that long-term consumption of
A. muricata is related to atypical Parkinsonian syndrome. The effect was later associated with annonacin which is an acetogenin presenting toxic activity against cultured dopaminergic mesencephalic neurons (
28). The determination of LD
50 was important for the choice of the dosage range used in this study; thus, it was decided to use dose 2,5%, 5% and 10% below the LD
50 (25, 50, and 100 mg/kg, respectively).
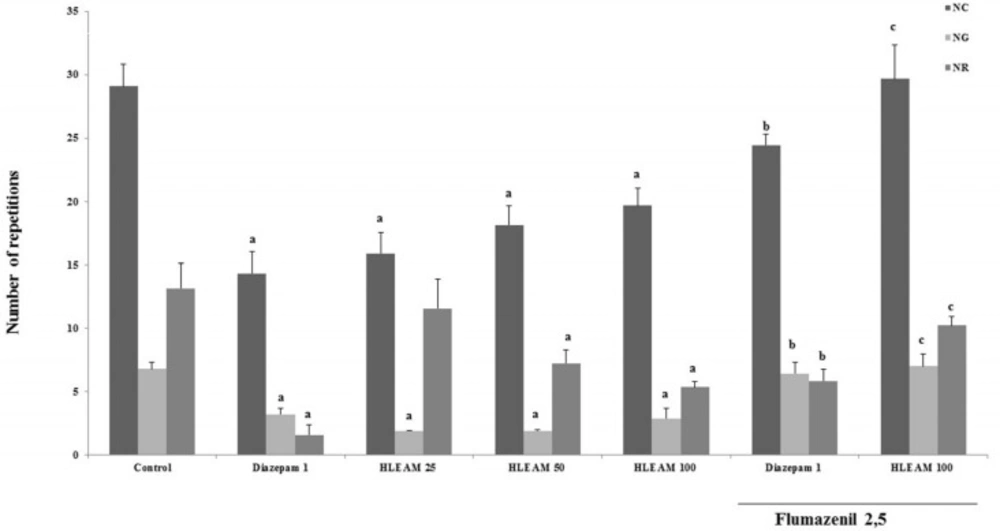
As the Hippocratic screening data showed that the HLEAM is endowed with central depressant activity, the animals treated with HLEAM were subjected to specific behavioral tests. In the open field test (
Figure 2), HLEAM 25, 50, and 100 mg/kg i.p., significantly reduced NC [(15.89 ± 1.65, 18.11 ± 1.54 and 19.67 ± 1.34, respectively); DF = 6, F (6.65) = 12.99,
P < 0.0001], NG [(1.88 ± 0.42, 1.88 ± 0.11 and 2.87 ± 0.84, respectively); DF = 6, F(6.57) = 12.20,
P < 0.0001] and NR [(11.56 ± 2.31, 7.22 ± 1.05 and 5.33 ± 0.47, respectively) DF = 6, F (6.56) = 8.13,
P < 0.0001], similar to diazepam (14.31 ± 1.71, 3.22 ± 0.49 and 1.55 ± 0.80, respectively) as compared to the respective controls (29.08 ± 1.73, 6.80 ± 0.50 and 13.13 ± 1.99). This suggests that the compound has sedative action, characterized by decreased spontaneous locomotion, possibly by decreasing the excitability of the central nervous system (
29).
Open Field Test – Effect of HLEAM without and with Flumazenil
It is known that both the nigrostriatal and mesolimbic dopaminergic systems play a crucial role in motor control; mesolimbic dopaminergic system, specially, is more closely linked to the control of locomotor activity (
30).
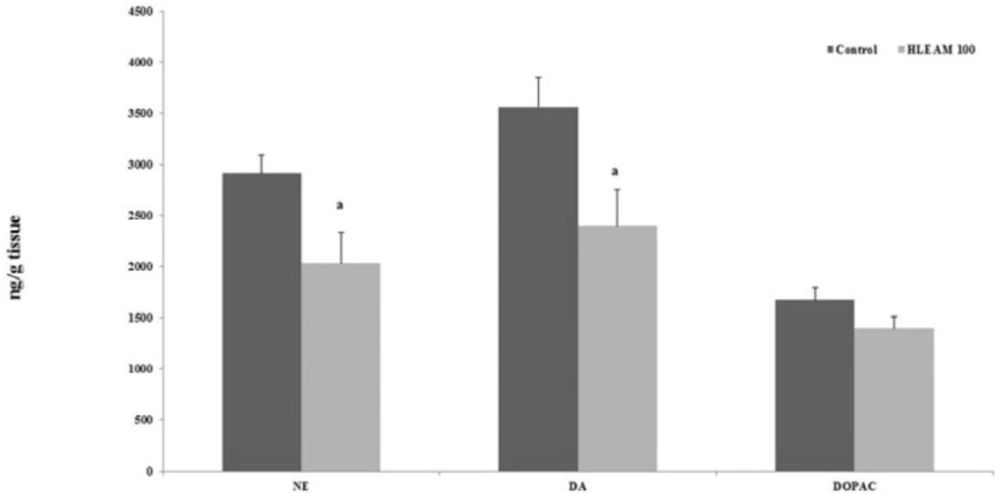
Thus, the sedative effect of HLEAM may be due to reduced levels of striatal dopamine (DA) and norepinephrine (NE), as evidenced by the HPLC test (
Figure 6). These catecholamines are abundantly present in the central nervous system, and modulate neuronal excitability (
31) and drugs such as reserpine, which lead to depletion of catecholamines that may cause sedation, catalepsy, or hypokinesia (
32).
The interaction of one or more components of HLEAM with the GABAergic system may be also responsible for the sedative effect observed with the extract. This hypothesis is supported by the fact that the pretreatment with Flumazenil (2.5 mg/kg, i.p. – benzodiazepine antagonist) increased NC, NG, and NR in diazepam (24.45 ± 0.85, 6.44 ± 0.85, and 5.84 ± 0.91, respectively) as well as HLEAM 100 (29.67 ± 2.67 and 7.00 ± 0.96, respectively) treated groups.
This result can be attributed to the presence of flavonoids identified in the present study (
Figure 1), indicating that these substances have neuroactive properties (
33) especially, quercetin and kaempferol, which were classified as anxiolytics constituents (
25).
Selvakumar (
34) had shown that quercetin reduced locomotor activity in the open field test suggesting that in the present study these compounds may be responsible for the anxiolytic and sedative effects.
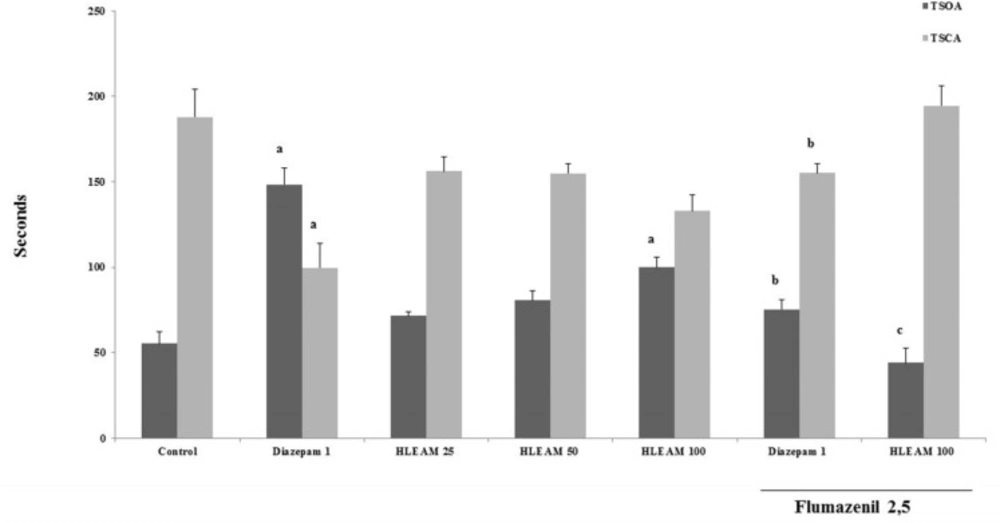
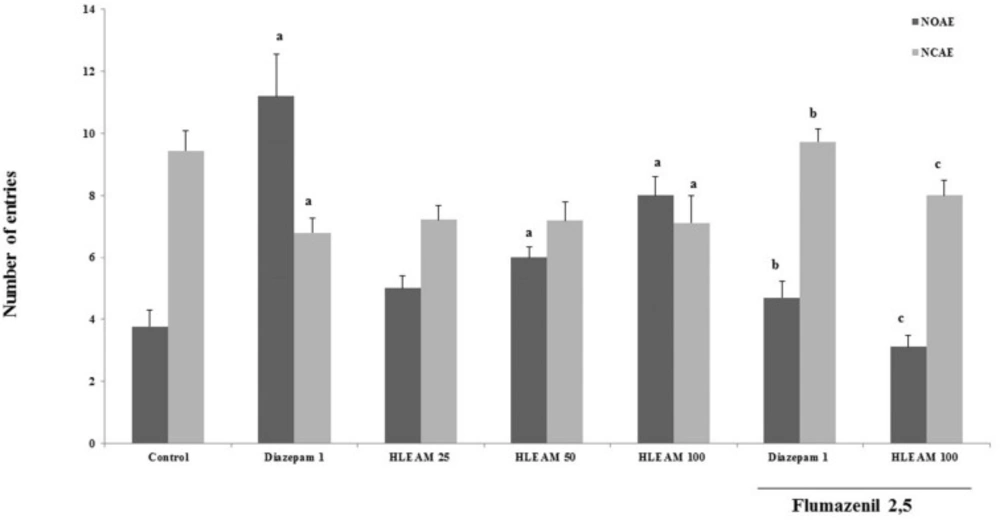
In the EPM test (
Figures 3 and
4), the mice were pre-treated with HLEAM 25, 50, and 100 mg/kg, i.p.; however, only the highest dose of the HLEAM significantly increased NEOA [(5.00 ± 0.40, 6.00 ± 0.33 and 6.00 ± 0.33, respectively) DF = 6, F (6.56) = 16.85,
P < 0.0001] and TSOA [(71.44 ± 2.66, 80.78 ± 5.52 and 100.0 ± 5.85, respectively) DF= 6, F (6.56) = 24.45,
P < 0.0001] and decreased TSCA [(156.2 ± 8.31, 155.0 ± 5.80 and 132.9 ± 9.27); DF = 6, F(6.56) = 8.58,
P < 0.0001], compared to controls (3.77 ± 0.54, 55.25 ± 7.22 and 187.9 ± 16.29, respectively). In the test, diazepam 1 mg/kg, significantly increased NEOA (11.20 ± 1.37) and TSOA (148.5 ± 9.80) and decreased NECA (6.79 ± 0.49) and TSCA (99.67 ± 14.32). Anxiolytic drugs used, especially the benzodiazepines, modulate GABA
A receptors (
35).
Elevated Plus Maze Test – Effect of HLEAM without and with Flumazenil on the time spent in the open and closed arms
Elevated Plus Maze Test – Effect of HLEAM without and with Flumazenil on the number of entries in the open and closed arms
The diazepam presents anxiolytic effect by increasing the number of entries (and time spent) in the open arms, reducing at the same time, the number of closed arm entries and time spent. Administration of HLEAM developed a pattern of effects similar to that observed with the diazepam. Elevated plus maze test is considered as an important experimental model for the study of drugs with anxiolytic effects of the benzodiazepine type (
36).
This observation may indicate that the anxiolytic-like effect of HLEAM was due to the interaction of one or more of the extract constituents with the GABA
A receptors. Wasowski (
37) showed that some types of flavones possess selective affinity for the benzodiazepine binding site on the GABA
A receptors in the brain. This finding confirms the study of Díaz-Vélez and Mora (
6) which interconnects the anxiolytic effects of
Annona muricata (0,5g/v.o.) to the GABAergic system.
To investigate the possibility that a lower dose tested by Diaz-Velez and Mora (
6), possibly modulates GABA
A receptors, the HLEAM 100 mg/kg was associated with 2.5 mg/kg flumazenil, a benzodiazepine antagonist with affinity for the GABA
A receptor (
38). Pretreatment with flumazenil partially reversed the effect of both, diazepam, and HLEAM in the elevated plus maze, by decreasing the NEOA (4.67 ± 0.52 and 3.11± 0.38, respectively) and TSOA (75.32 ± 5.86 and 44.14 ± 8.31, respectively) and increasing the NECA (9.73 ± 0.42 and 8.00 ± 0.5, respectively), and TSCA (155.1 ± 5.79 and 194.4 ± 12.15, respectively), significantly in relation to the group treated with HLEAM or diazepam alone.
Benzodiazepines have several effects related to modulation of GABA
A receptor, such as sedation, reduced muscle tone, and diminished motor coordination; this latter is a common feature of many neurological disorders and pharmacological intoxication (
39,
40). The benzodiazepines compromise the execution of activities that require reflexes and motor control by inhibiting polysynaptic reflexes and internuncial transmission, and in high doses can depress transmission at the skeletal-neuromuscular junction (
41). The rota-rod test is used to evaluate the effect of drugs on motor coordination, allowing detection of neurological disorders including ataxia, sedation, and muscle relaxation, in addition to neurotoxicity (
42,
43).
In the rota-rod test, HLEAM (at all doses) had no effect on motor coordination of the animals (data not shown). This observation is important because despite its sedative and anxiolytic effect, the HLEAM, at the doses tested, did not promotes muscle either relaxation or loss of reflexes, the unwanted effects commonly seen with benzodiazepine drugs.
Once the HLEAM presented central depressant effect, the anticonvulsant effect by pentylenetetrazole-induced seizures model was investigated. The mechanisms of seizure, activation, propagation, and maintenance are poorly understood. Seizures can be characterized as clinical manifestations resulting from abnormal neuronal discharges, producing over-excitation of the neurons, and also occur by imbalances between the mechanisms of inhibitory and excitatory neurotransmission (
44).
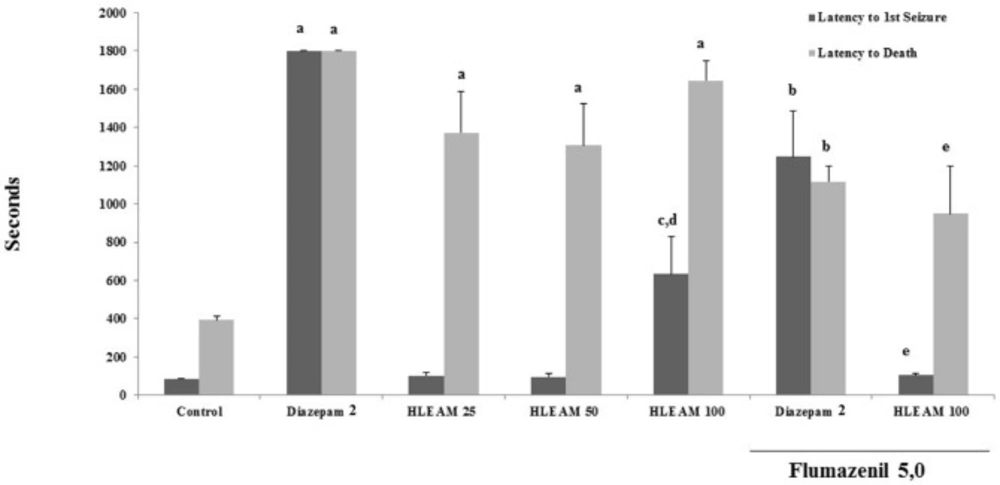
The mice were pre-treated with HLEAM 25, 50, and 100 mg/kg i.p.; however, only the highest dose of HLEAM showed significant increased latency until the first seizure [(102.3 ± 17.59, 97.44 ± 14.03, and 633.9 ± 194.0, respectively); DF = 6, F (6.56) = 59.91,
P < 0.0001], when compared to the control (87.00 ± 1.46). On the other hand, HLEAM 25, 50 and 100 mg/kg increased significantly the death latency parameter [(1374 ± 215.6, 1307 ± 215.6 and 1646 ± 101.6, respectively); DF = 6, F (6.56) = 8.39,
P < 0.0001], as well as diazepam 2 mg/kg, i.p. (1800 ± 0.0) compared to the control (393.7 ± 19.91) (
Figure 5).
Pentylenetetrazole-induced seizures test – Effect of HLEAM without and with Flumazenil
According to Oviedo
et al. (
7)
A. muricata did not confer protection against electroshock or pentylenetetrazol-induced seizures. The difference may be related to dosages used or to absorption differences due to the route of administration (in the Oviedo
et al. study, the extract was administered orally). The effect observed in this reasearch may be due to the action of the extract on the GABAergic system. When associated with flumazenil 5 mg/kg, HLEAM 100 mg/kg decreased the first convulsion and death latency of the animals (105.8 ± 9.07 and 949.8 ± 248.6, respectively) compared to the group treated with HLEAM 100 mg/kg only. Similarly, the association flumazenil 5 mg/kg and diazepam 1 mg/kg decreased the first convulsion and death latency of the animals (1248 ± 238.3 and 1117 ± 80.91, respectively), when compared to the group treated with diazepam 1 mg/kg alone.
In
Figure 6 the administration of HLEAM caused a significant decrease in the levels of NE [2032 ± 297.3; DF = 4, DF (3.3) = 2.74, P = 0.044] and DA [2396 ± 350.8; DF = 4, F (3.3) = 1.48,
P = 0.042], compared to their respective controls (2912 ± 179.3 and 3559 ± 287.7). The concentration of DOPAC [1397 ± 115.5; DF = 4, F (3.3) = 1.21,
P = 0.152] did not change with HLEAM administration, compared to the control group (1673 ± 122.3). In the present study, HLEAM showed a neuroprotective effect, although less than the diazepam in PTZ-induced seizure test. Shouse
et al. (
45) asserted that the increase in noradrenaline and serotonin, (but not dopamine), would be accompanied by electroencephalogram changes, due to intermittent neuronal discharges in the amygdala, (and/or the locus coeruleus), and/or widespread throughout the brain cortex during the seizure.
High Performance Liquid Chromatography (HPLC) – Effect of HLEAM treatment on striatic monoamine and metabolite contents
Thus, the neuroprotective effect of HLEAM may be also due to the reduction in noradrenaline concentration as was evidenced by the high performance liquid chromatography study.