Cell culture
MKN-45 (nucleolin positive) and HDF (nucleolin negative) cell lines were initially obtained from Pasteur Institute (Tehran, Iran). MKN-45 cells and HDF cells were grown in RPMI-1640 medium and Dulbecco’s modified Eagle’s medium (DMEM/F12), respectively at 37 °C in humidified air containing 5% CO2. The culture media were supplemented with 10% fetal bovine serum (FBS), 100 IU/mL penicillin, and 100 µg/mL streptomycin. The cell culture materials mentioned above were acquired from Gibco Company (Karlsruhe, Germany).
Oligonucleotidesand chemicals
All of the oligonucleotides were synthesized by Bioneer (Korea). The sequences were as follows: 3′-thiolated AS1411 (aptNCL): 5ʹ -GGT GGT GGT GGT TGT GGT GGT GGT GGA AAA A- 3ʹ (SH). Control aptamer: 5ʹ- GTA CAT TCT AGA TAG CC -3ʹ. miRNA let-7d 5p: 5ʹ- AGA GGU AGU AGG UUG CAU AGU U- 3ʹ. 3ʹ aminated miRNA let-7d 3p: 5ʹ -CUA UAC GAC CUG CUG CCU UUC U- 3ʹ (NH2). AS1411-stick: 5ʹ -GGT GGT GGT GGT TGT GGT GGT GGT GGA AGG CTA TCT AGA ATG TAC- 3ʹ. miRNA let-7d 3p-stick: 5ʹ -CUA UAC GAC CUG CUG CCU UUC UGU ACA UUC UAG AUA GCC UU- 3ʹ and miRNA mimic negative control (#SMC-2001, Bioneer, Korea).
According to the company guidelines, thiolated sequences were initially subjected to the reduction of SH terminal group and then immediately used in the following reactions. In the primary step, aptamer sequences were exposed to a short denaturation–renaturation stage to form G-quartet structure, as they were heated at 88 °C for 10 min and then cooled to room temperature (RT) for 15 min (
22).
Chemicals were PEGylated SMCC crosslinker (Thermo Fisher Scientific, USA), Dithiothreitol (DTT) (Sigma, USA), Lipofectamine 2000 (Sigma, Germany), DMSO (Gibco, Germany), Acrylamide (Sumchun, China), TEMED (Bio Rad, Germany), and DNA ladder 10bp (GeneOn, Germany).
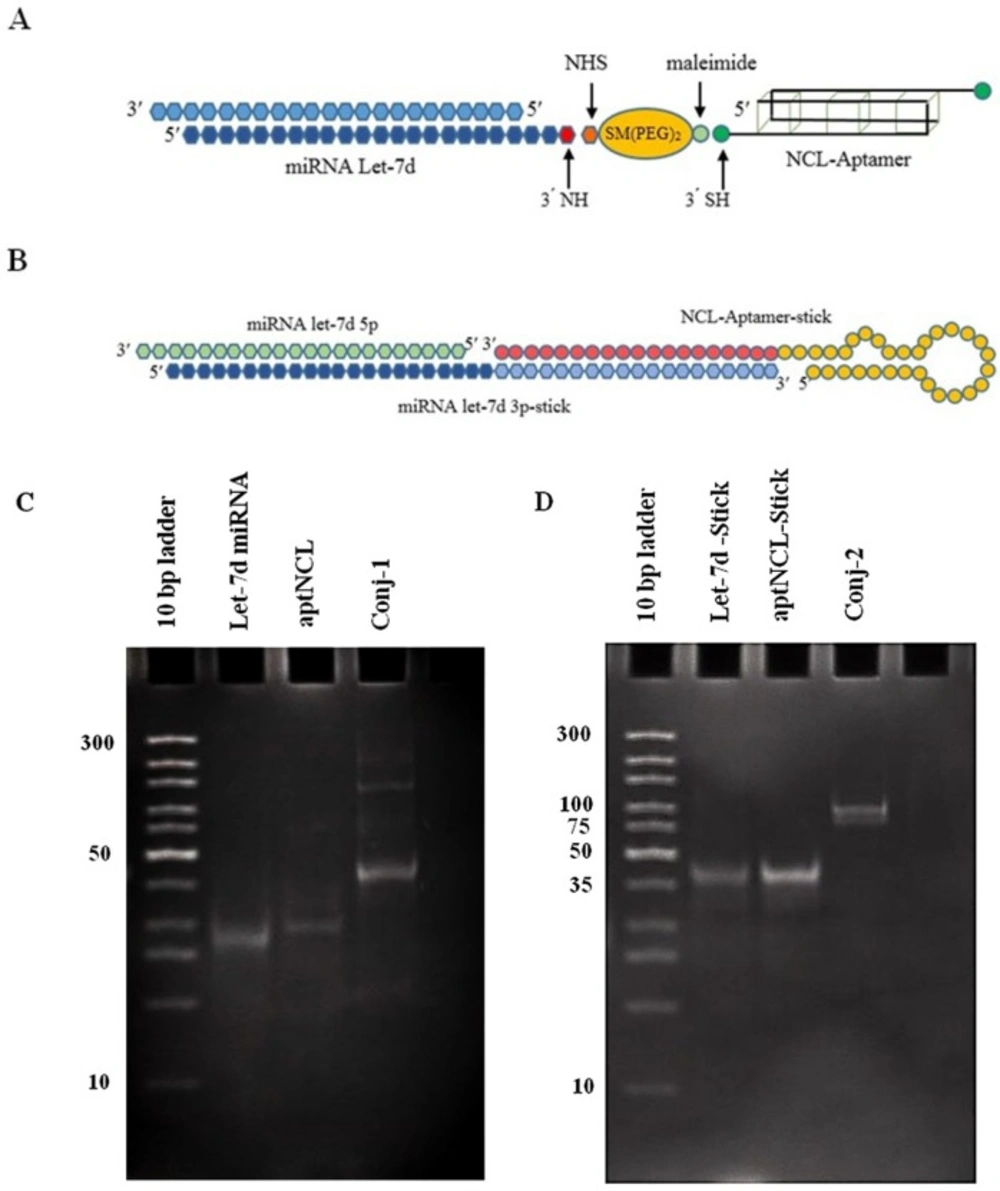
The aptNCL-miRNA conjugate was consisted of two entities, aptNCL and miRNA let-7d, which was linked together by two different methods:
Preparation of aptNCL -miRNA let-7d conjugate using SM (PEG)2 cross linker
As the first step, disulfide bonds in Thiol-modified aptNCL were reduced to SH group (SH-apt). Briefly, 100 µL of thiolated aptamer solution (6 nmol) was mixed with 100 µL of 1M DTT solution (pH 6) and incubated at RT for 45 min followed by 37 °C for 15 min. To eliminate excess DTT, the solution was extracted three times by the same volume of Ethyl acetate.
At the second step, the amine-modified miRNA let-7d duplex was incubated with a 50-fold molar excess of SM (PEG)2 in PBS (pH 7.2) containing 3mM EDTA as the conjugation buffer at 37 °C for 1 h, then at 4 °C overnight. Excess SM (PEG) 2 was removed using Amicon Ultra 0.5 mL Centrifugal Filter (cut off: 3kD; Merck Millipore, Germany). The sample was centrifuged and equilibrated with the conjugation buffer. Finally, the column was reversed and centrifuged to collect the purified sample. At the last step, purified miRNA let-7d-SM (PEG)2 and aptNCL-SH were mixed together in equal molar ratio at RT for 1 h, then an overnight at 4 °C. The conjugate formation was confirmed by poly acrylamide gel electrophoresis (PAGE, 15%) and the construct stability was evaluated in human serum.
Preparation of aptNCL -miRNA let-7d conjugate using sticky ends
Equimolar ratio of aptamer-stick, miRNA let-7d 5p, and miRNA let-7d 3p-stick were separately dissolved in conjugation buffer (pH 7.4) and heated at 95 °C for 2 min, then cooled at 37°C for 15 min. Then, immediately were added to each other and incubated at 30 °C for 30 min followed by an adjustment step at RT for 30 min. The conjugation efficacy was assessed by PAGE 15%, and the stability of the conjugate was examined in human serum.
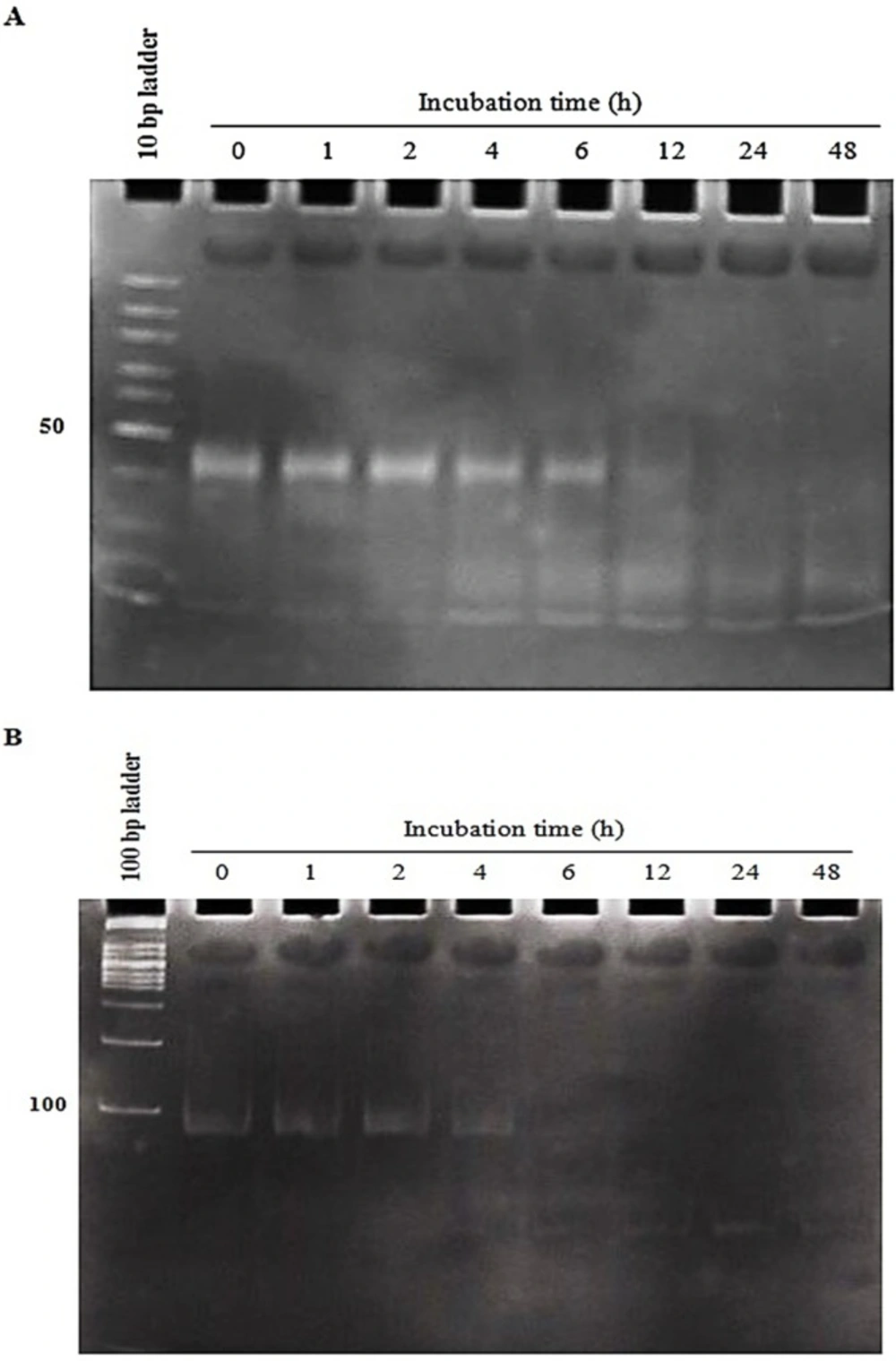
Evaluation of aptNCL -miRNA let-7d stability in human serum
To evaluate the serum stability of both conjugates, 4 µM of the conjugates were subjected to 80% human serum (Type AB
+) from 1 to 48 h. At the indicated time intervals (0, 1, 2, 4, 6, 12, 24 and 48h), 8 µL (32 pmol) of each sample was extracted and incubated at 37 °C for 2 h with 1 µL proteinase K solution (600 mAU/mL) (Sigma, Germany) to eliminate serum proteins interference in electrophoresis. Then, 9 µL from 1X Tris
-Borate
-EDTA (TBE) and 3 µL of gel loading buffer (GeneOn, Germany) were added and kept at -80 °C. Finally, all samples were analyzed by 15% PAGE. The gel was stained by SYBR Green DNA safe stain (Sinaclone, Iran) (
25).
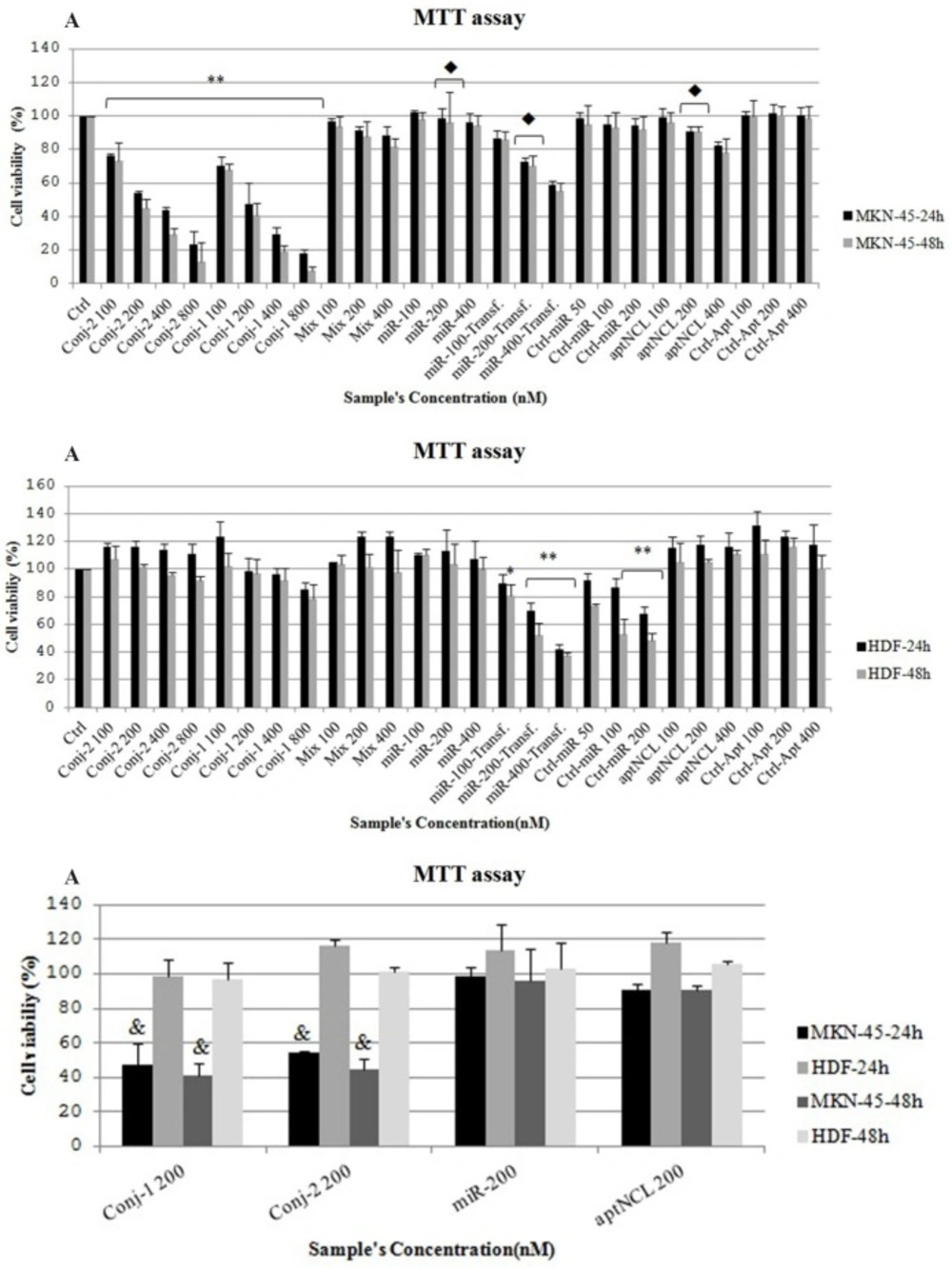
Cell proliferation assay
Cell proliferation was assessed by using 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay kit (BioBasic, Canada). Briefly, MKN-45 and HDF cells were seeded at 7,000 cells per well (96-well plates) in their respective media. After 24 h of seeding, and following 4 h of starvation using the serum-free media, the cells were treated with the conjugates , aptNCL, control aptamer, mixture of aptNCL plus miRNA let-7d, or transfected with miRNA let-7d , miRNA mimic negative control for 8h and incubated for 24 and 48 h before MTT assay, separately. MTT solution (100 µL/well, 5 mg/mL) was added to each well and then incubated for 4 h at 37 °C. After removing the medium, 100 µL of DMSO was added in each well and the plate was shaken for 10 min at 100 rpm for dissolving the formazan crystals. The absorbance of each well was measured by a microplate reader (Stat Fax-2100, Awareness, USA). The experiments were done in triplicate. The results of each treatment were normalized with the control medium and relative proliferation activity was determined as a percentage of the control (activity of control was considered as 100). miRNA let-7d and miRNA mimic negative control were transfected using Lipofectamine 2000 according to the manufacturer’s protocol. The half maximal inhibitory concentration (IC50) of the conjugate-1 and conjugate-2 was determined for 48 h (
26).
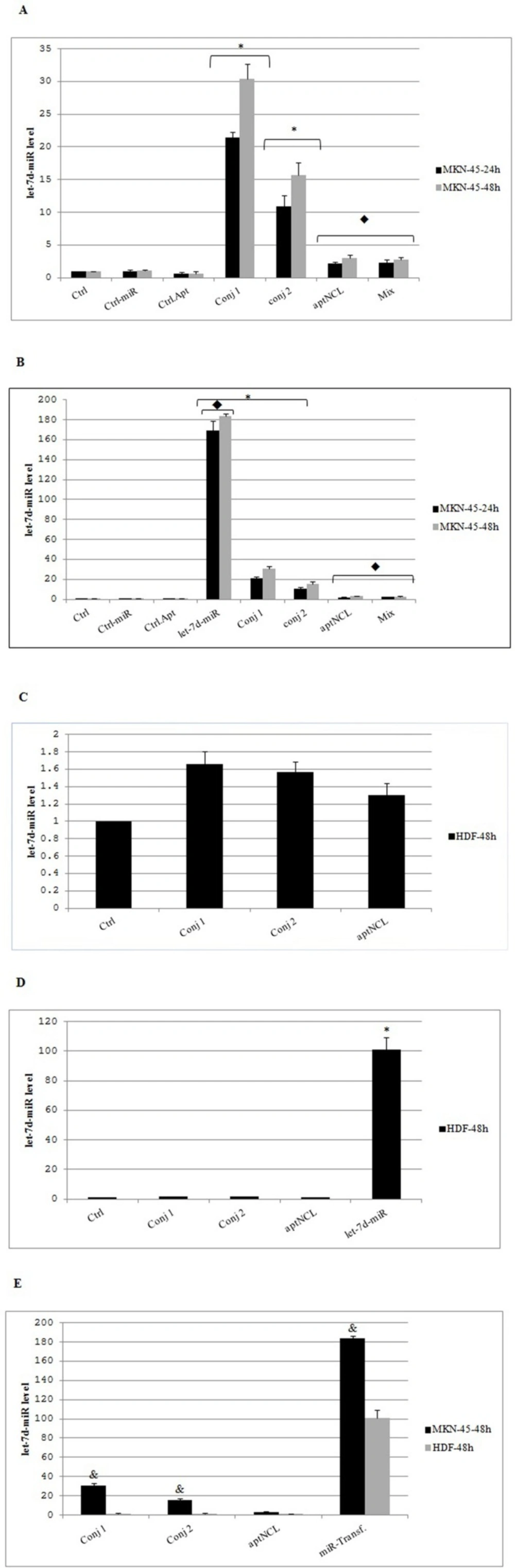
miRNA let-7d quantification using quantitative RT-PCR
MKN-45 and HDF cells were seeded at 40,000 cells per well (24-well plates) in their relevant media. After 24 h of seeding, and following 4 h of starvation using the serum-free media, the cells were treated with the conjugates (at IC50 concentrations), aptNCL, control aptamer, and mixture of aptNCL plus miRNA let-7d (200nM) or transfected with let-7d miRNA and control-miRNA (200nM) and incubated for 24 and 48 h, separately. In two time point of 24 h and 48h, the relative expression of miRNA let-7d was detected by quantitative RT-PCR technique. Briefly, RNAs were recovered by 1mL of RiboEx RNA extraction kit (Gene all, Korea) according to the manufacturer’s instructions. To evaluate the intracellular levels of the let-7d miRNA (target gene) or U6 RNA (housekeeping control gene) (Cat #MS00033740, Qiagen, Germany), 500 ng of total RNA was reverse transcribed by using miScript II Reverse Transcription cDNA synthesis kit (Qiagen, Germany) in a final volume of 20 µL consistent with manufacturer’s protocol: 60 min 37 °C then 5 min 95 °C by Mastercycler gradient (Eppendorf, Germany), then, amplification was done using the miScript-SYBR Green PCR Kit and specific miScript Primer Assay (Cat #MS00003136, Qiagen, Germany) and analyzed using the StepOneTM cycler real-time PCR system (Applied Biosystems, USA). The ΔΔCT method was used for relative quantization of miRNA levels. Each sample was run in triplicate.
Statistical Analysis
Data are presented as mean ± SD. Inter-group comparisons were performed using the one-way analysis of variance (ANOVA) followed by post hoc Tukey’s test. In each group, Paired t-test was used to compare between 24h and 48 h. For all tests, P < 0.05 was considered as statistically significant. The data were analyzed using SPSS software version 22.