Chemicals
Skimmed milk, Folin-Ciocalteu reagent (FCR), culture media and buffer components, and also surfactants were purchased from Merck (Darmstadt, Germany). Casein, phenylmethylsulfonyl fluoride (PMSF), and tyrosine were provided from Sigma-Aldrich (St. Louis, MO, USA). Other chemicals were of analytical grade and were commercially available.
The screening of halophilic bacteria for the extracellular protease activity and identification of the selected strain
To isolate the halophilic bacteria, the samples were collected aseptically from different salty districts of Iran. Isolation of halophilic bacteria using salt enrichment was done using the modified Luria-Bertani (LB) agar medium containing (g L-1 distilled water): yeast extract, 5; peptone, 10; agar, 15; and NaCl, 200. pH of the medium was adjusted at 7.5 and sterilized by autoclaving (121 °C and 15 psi for 20 min). Afterwards, 1 g soil was suspended in 1 mL sterile phosphate buffer saline (PBS) containing 20% NaCl, and after vortexing, 500 μL of suspension (or water samples) was spread over the LB agar plates. After incubating at 37 °C for 48 h, isolation was completed by serial dilution and plated on the LB agar medium. The colonies were often streaked on the LB agar medium for purification and stored on the modified agar plates at 4 °C. Stock cultures of the pure bacterial colonies were maintained as a glycerol stock (the modified LB broth medium containing 20% w/v NaCl and 15% v/v glycerol) at −85 °C.
To screen the protease-producing colonies, the purified bacteria were transferred to the skimmed milk agar plates (1% (w/v) skimmed milk powder and 20% (w/v) NaCl). The colonies with the proteolytic activity showed clear zone. The isolated strains with the maximal protease activity, as measured by the ratio of hydrolytic zone diameter (mm) to the colony diameter (mm), was subjected to screen for the protease production in the modified LB broth medium (containing 20% NaCl). The isolate with the highest proteolytic activity in the modified LB broth medium was subjected for subsequent studies.
To identify the strain, the selected pure strain was streaked on the modified LB agar plate as described above and incubated for 48 h at 37 °C. The applied morphological tests were gram staining, checking the shape and color of the colony, and spore-forming ability. The biochemical and physiological characteristics were further determined using a LB 48 h-old culture of the purified isolate and included catalase and oxidase, motility, Voges-Proskauer, and methyl red tests, growth at different temperatures, pH values, and NaCl concentrations, the fermentation of some sugars, hydrolysis of starch, gelatine, casein, Tween 80, nitrate reduction, and H2S and indole production. For molecular characterization, the bacterial genomic DNA of the isolate was extracted by Genomic DNA Extraction Kit (Bioneer, Korea) under the manufacturer’s protocol. A pair of 16S rRNA universal primers, including 27F as a forward primer (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1525R as a reverse primer (5́ -TTCCTCCACTAGGTCGG-3′) (15), were used to amplify a ~1.5 kb fragment of 16S rRNA gene using polymerase chain reaction (PCR) was performed in a reaction volume of 100 µL containing PCR buffer with 1.5 mM MgCl2, 200 µM dNTPs, Taq DNA polymerase (5.0 U), 1.0 µM each primer, and DNA template (50 ng). The PCR amplification was programmed as follows: (i) initial denaturation at 94 °C for 5 min; (ii) 30 cycles of denaturation at 94 °C for 1 min, annealing at 56 ºC for 1 min and extension at 72 ºC for 1.5 min; and (iii) final elongation at 72 °C for 10 min. Following the visualization of the amplified DNA using gel electrophoresis, DNA fragment of ~ 1.4 kb was eluted from gel using a QIAGEN gel extraction kit. The extracted fragment was employed as a template for amplifying forward and reverse strands using the same primers, and separate sequencing of the PCR products. The phylogenetic neighbors were determined by BLASTn (NCBI, http://ncbi.nlm.nih.gov/BLAST/) against GenBank® database at the NCBI server. As a result, strain SR-081 Halo (sampled from Golestan Salt Lake, Iran (37°24′3′′N 54°38′44′′E)) was designated as Salicola marasensis (S. marasensis) and deposited in NCBI with the accession number of KF859984 (http//www.ncbi.nlm.nih.org/ available Jan 18, 2018). Then, the 16S rDNA sequences of the close relatives of the isolate were retrieved from the GenBank® database and the phylogenetic tree was constructed by the neighbor-joining method. The significance of the junctions was stablished using the bootstrap method (1000 replicates).
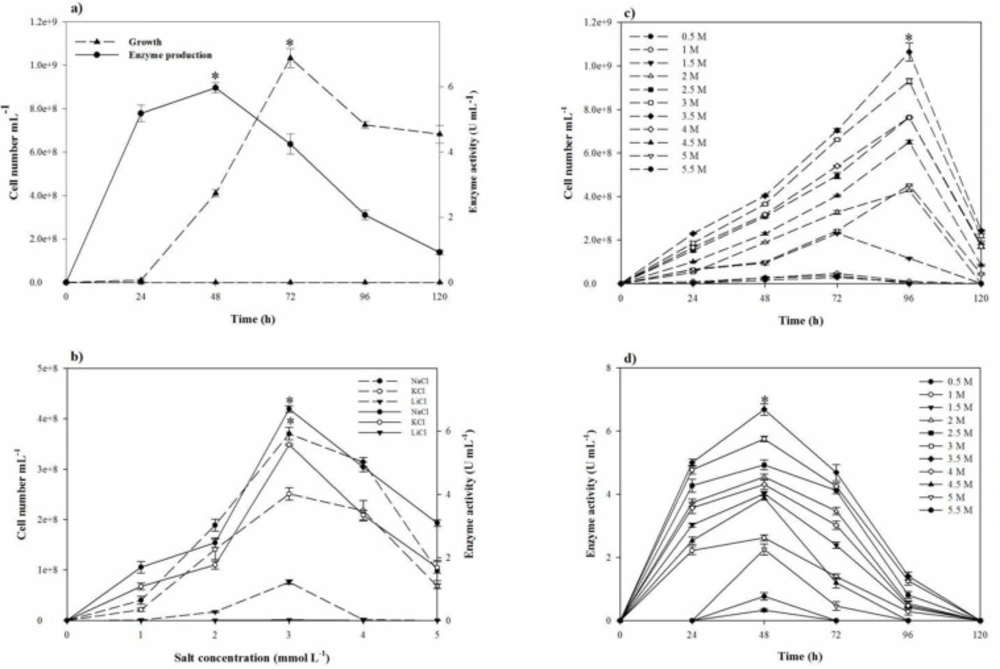
Media, growth behavior, and the production ofproteolytic extract by S. marasensis
Growth and protease production of
S. marasensis were studied in the minimal basal medium (
16) consisting of (g L
−1): yeast extract, 5; MgCl
2.6H
2O, 20; K
2SO
4, 5; CaCl
2.2H
2O, 0.1; and NaCl, 200. Fifty mL of the basal medium were taken in 250 mL Erlenmeyer flasks and pH was adjusted to 7.4 before being autoclaved at 121 °C and 15 psi for 20 min. The flasks were inoculated with 1 mL of a two-day-old mother culture of
S. marasensis (OD
600 ~ 1) and incubated at 37 °C with continuous shaking at 150 rpm for 120 h. The samples were withdrawn periodically at the 24-h intervals and analyzed for the protease production and the growth estimation. Bacterial growth was estimated by the cell number determination. To oversee the proteolytic activity, the withdrawn samples were centrifuged at 8,000×g for 10 min and the cell-free supernatant was assayed for the extracellular proteolytic activity.
Protease assay and protein determination
Proteolytic activity was determined toward casein following the method described by Kunitz and Gen (
17) with some modifications. Briefly, 0.5 mL of casein (0.5% (w/v) prepared in 50 mM phosphate buffer, pH 8.0, containing 20% (w/v) NaCl was added to 1 mL (~ 5 U) of the enzyme sample to start the reaction. The mixture was immediately incubated at 40 °C for 15 min, and the reaction was stopped by adding 0.5 mL of 60% TCA. After 5 min standing at room temperature, the precipitate was centrifuged at 18,000×g for 10 min to be removed and the supernatant was used to estimate the free tyrosine amount. One mL of Na
2CO
3 (0.5 M, to naturalize the supernatant) and 250 µL of the FCR (1 N) were added to 0.5 mL of the supernatant. Followed by keeping the mixture at 37 °C for 15 min, the absorbance was recorded with an UV/Vis spectrophotometer at 660 nm against the corresponding blank. As a blank, the enzyme was inactivated at the beginning of the incubation period. The proteolytic activity of the samples was measured based on the tyrosine standard curve, and one unit of proteolytic activity was expressed as the enzyme quantity, releasing 1 µmol of tyrosine min
−1 mL
−1 under the standard assay conditions. The standard curve was made by 0–1000 µmol mL
−1 of tyrosine. All analyses were carried out in triplicate; the results were taken in the means of three independent determinations. In addition, the protein concentration was determined by the dye binding assay suggested by Bradford (
18), using bovine serum albumin (BSA) to prepare the standard solution range.
Medium optimization for maximum protease production by S. marasensis
Preliminary study
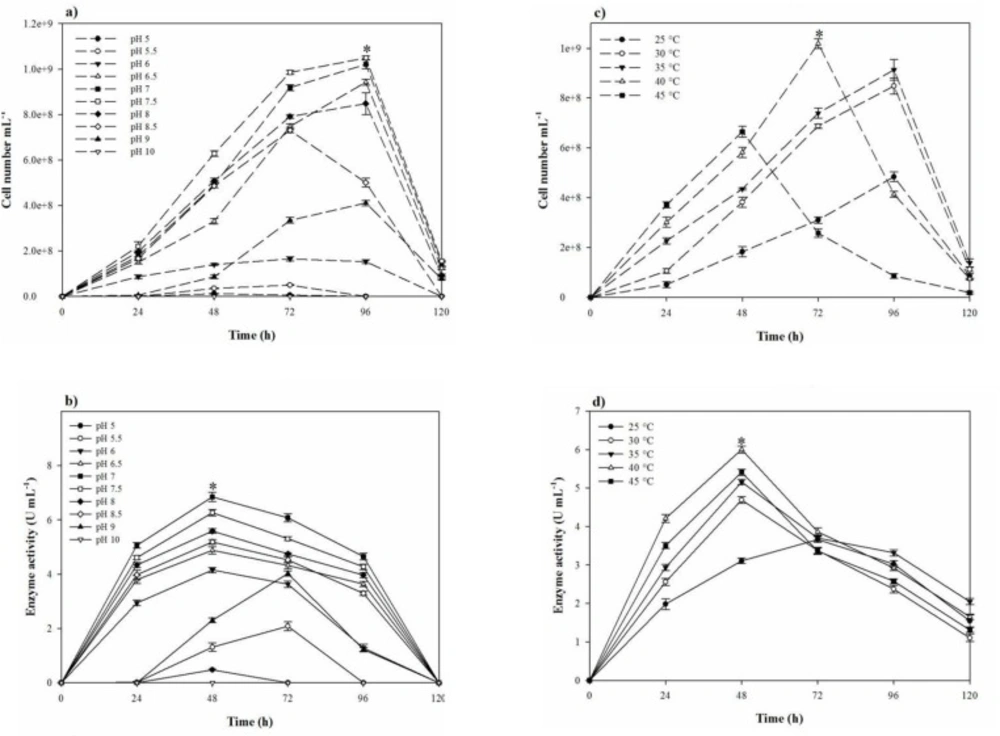
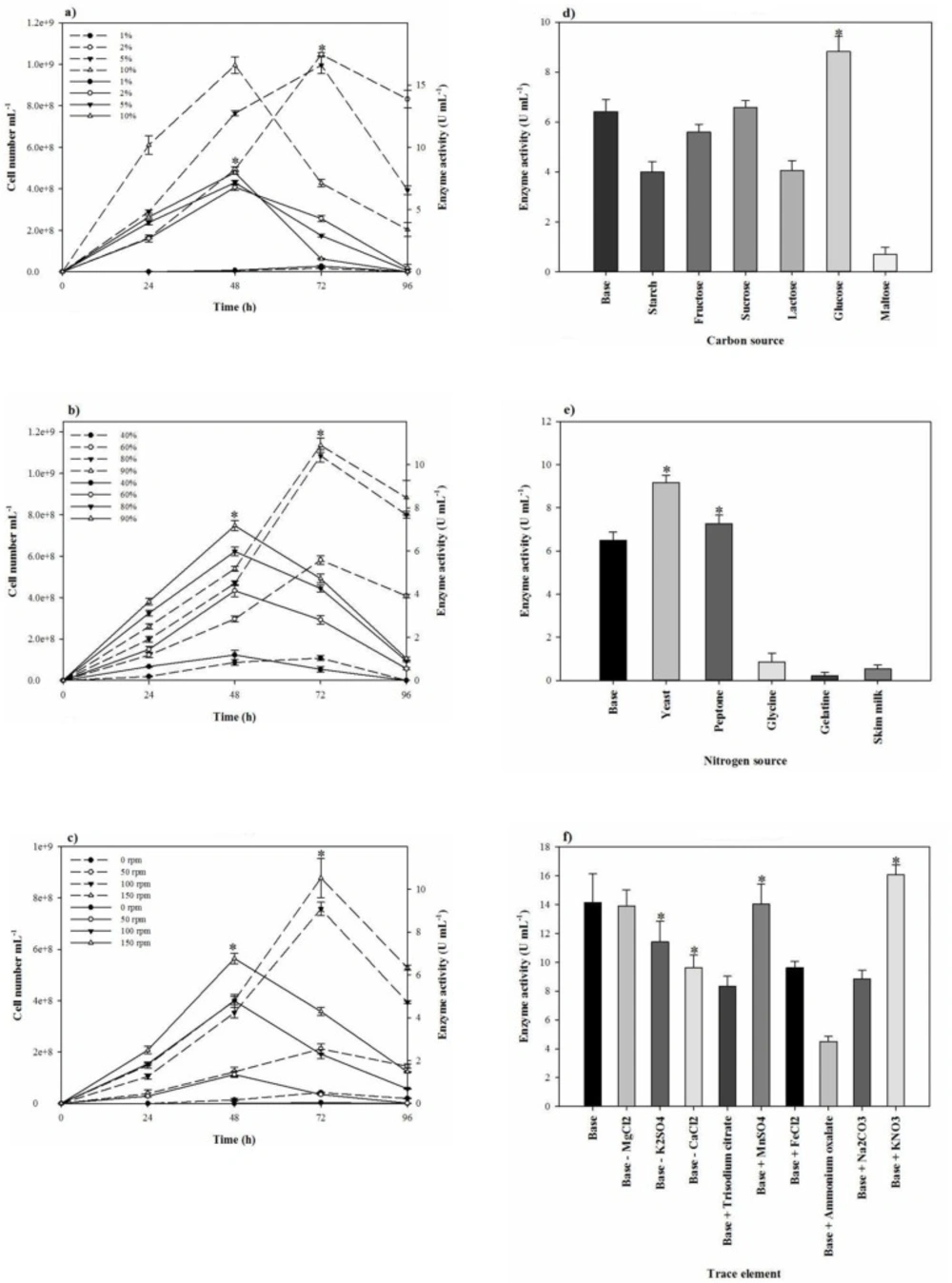
Considering that the broad applications of halophilic proteases demand efficient production, one-variable-at-a-time strategy was applied to early screen of the most significant items needed to increase the growth and proteolytic activity. Therefore, the conditional tests, including temperature (25–45 °C), pH (5.0–10.0), inoculum size (1–10%), agitation speed (50–150 rpm), fermentation period (24–120 h), and aeration (40–90%, according to the total volume of Erlenmeyer flask occupied by air) were investigated. To find the most effective nutritional sources on the growth and proteolytic activity, several carbon sources (starch, fructose, sucrose, lactose, glucose, and maltose), nitrogen sources (yeast extract, peptone, glycine, gelatin, ammonium oxalate, potassium nitrate, and skimmed milk powder), trace elements (MgCl2.6H2O, K2SO4, CaCl2.2H2O, MnSO4, FeCl2, KNO3, trisodium citrate, and Na2CO3), and salts (NaCl, KCl, and LiCl) were implemented. For evaluating the effects of the nitrogen sources, the additive sources were applied at 0.5% (w/v) concentration in addition to 0.5% (w/v) yeast extract already present in the basal medium. Also all the carbon sources were added at 1% (w/v) level. The effect of a variety of inorganic elements on the proteolytic activity also was checked using individually removal of MgCl2.6H2O, K2SO4, and CaCl2.2H2O from the basal medium and individually addition of MnSO4, FeCl2, KNO3, trisodium citrate, and Na2CO3 to the basal medium at the final concentration of 0.05, 0.005, and 0.5% (w/v), respectively. Similarly, the effect of salts (NaCl, KCl, and LiCl at the concentrations of 0–30%) on the strain growth and proteolytic activity was evaluated.
Experimental design
The screening design: Taking the above factors with an effect on the proteolytic activity in consideration, the fractional factorial design (FFD) was adopted to identify the items having the most significant effects. In this design, a resolution III 2
(7-4) FFD was employed to reach the main effects of each factor and clarification of two-factor interactions. The effect of seven factors on the proteolytic activity at two levels (g L
−1) (based on the literature) of low (−1) and high (+1) were investigated after 48 h of incubation at 40 ºC and pH 7.0. The levels were included peptone (X
1; 0.1–1.0), glucose (X
2; 0.5–2.0), MgCl
2.6H
2O (X
3; 0.1–1.0), K
2SO
4 (X
4; 0.1–1.0), CaCl
2.2H
2O (X
5; 0.05–0.50), KNO
3 (X
6; 0.1–1.0), and MnSO
4 (X
7; 0.05–0.25). The matrix covering 11 experiments (3 were center point runs to show lack of fit) has been shown in
Table 1. The design was made and analyzed using statistical software package, Design-Expert (version 7.0.0; Stat-Ease; Inc., Minneapolis, Minnesota, USA). The model equation for 2
7-4 design was:
Y = β0 + β1 X1 + β2 X2 + … +β7 X7
where Y is the predicted response (proteolytic activity), β0 is the intercept, β1 is the coefficient for X1, and X1 is the first independent variable that explains the variance in Y. Statistical analysis of data with the aid of half-normal percentage probability plot and evaluation by analysis of variance (ANOVA) calculated p-value. Factors with p-value below 0.05 were considered to influence the proteolytic activity significantly and the model terms were selected or rejected based on the significance. All the experiments were performed in triplicate and the average of responses was attended.
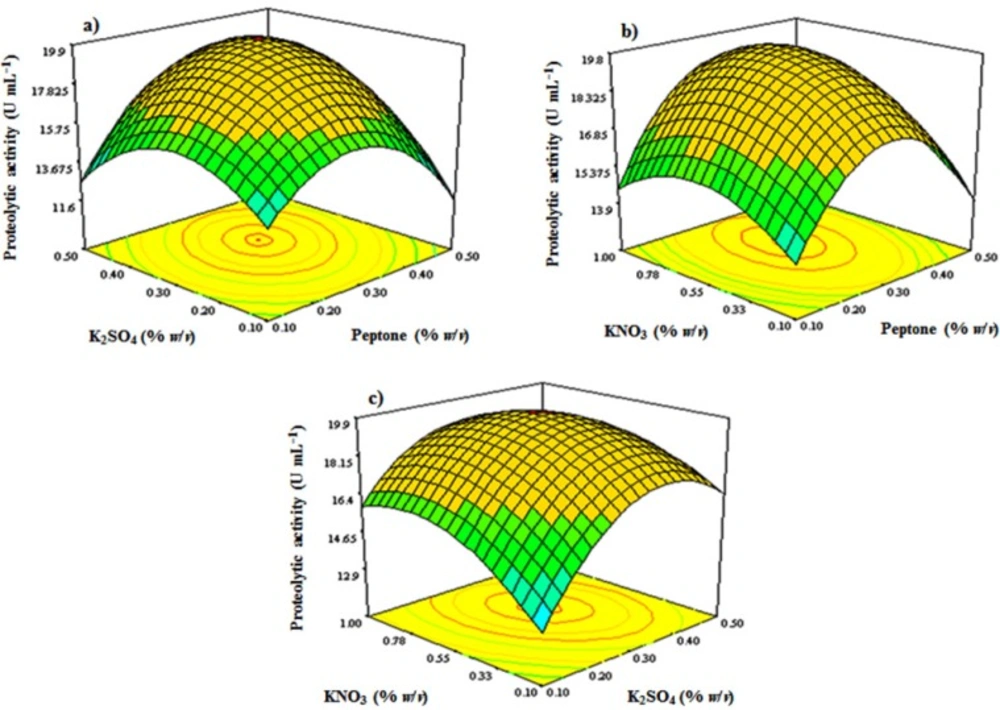
The optimization design: The selected significant elements from the preliminary screening study and FFD were subjected to the Box-Behnken design (BBD) (
19,
20) to optimize the proteolytic activity of
S. marasensis. Based on the screening results, peptone (X
1), K
2SO
4 (X
4), and KNO
3 (X
6) were fed to the Box-Behnken design to be optimized and each variable was evaluated at three experimental levels coded as −1, 0, and +1. The variables were checked at three levels (g L
−1): X
1 (−1: 1.0; 0: 3.0, and +1: 5.0), X
4 (−1: 1.0, 0: 3.0, and +1: 5.0), and X
6 (−1: 1.0, 0: 5.5, and +1: 10.0). All the variables were taken at a central coded value considered as zero. As a result, a total of 17 trials, including 12 factorial points and 5 repeats at the center point for estimation of the pure error sum of squares were performed (
Table 2). This design was applied using the Design-Expert® software. On completion of runs, the average maximal proteolytic activity yield was taken as the response (Y) and a second order polynomial equation was then fitted to the data by multiple regression procedure. For three-factors, a second-order polynomial equation that predicted the proteolytic activity was in the following form:
Y = β0 + β1 X1 + β2 X2 + β3 X3 + β12 X1 X2 + β13 X1 X3 + β23 X2 X3 + β11 X12 + β22 X22 + β33 X32
| Trial NO. | Variables
| Response (U mL−1) |
|---|
| X1 | X2 | X3 | X4 | X5 | X6 | X7 |
|---|
| 1 | −1 | −1 | −1 | +1 | +1 | +1 | −1 | 10.4 ± 0.3 |
| 2 | +1 | −1 | −1 | −1 | −1 | +1 | +1 | 11.4 ± 0.4 |
| 3 | −1 | +1 | −1 | −1 | +1 | −1 | +1 | 10.7 ± 0.3 |
| 4 | +1 | +1 | −1 | +1 | −1 | −1 | −1 | 7.7 ± 0.3 |
| 5 | −1 | −1 | +1 | +1 | −1 | −1 | +1 | 8.2 ± 0.2 |
| 6 | +1 | −1 | +1 | −1 | +1 | −1 | −1 | 8.9 ± 0.2 |
| 7 | −1 | +1 | +1 | −1 | −1 | +1 | −1 | 12.0 ± 0.6 |
| 8 | +1 | +1 | +1 | +1 | +1 | +1 | +1 | 9.1 ± 0.3 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9.5 ± 0.3 |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10.5 ± 0.4 |
| 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10.9 ± 0.4 |
where Y is the predicted response, β0 is the intercept, β1-β3 are the regression coefficients, β11-β33 are the squared effects, and X1-X3 are the independent variables. The fitness of the second order polynomial equation was evaluated by multiple correlation coefficient (R2) and adjusted R2. The responses under different combinations defined by design were analyzed using ANOVA to estimate the statistical parameters. Then, the response surface plots were constructed to study the interaction among factors. To validate the response surface model and evaluate the precision of the model, a random set of the optimized checkpoints experiments was set up according to the conditions predicted by the model. The experiments were conducted both within and outside the design space.
| Trial NO. | Variable
| Response (U mL−1) |
|---|
| Peptone | K2SO4 | KNO3 |
|---|
| 1 | −1 | −1 | 0 | 13.9 ± 0.6 |
| 2 | +1 | −1 | 0 | 10.5 ± 0.5 |
| 3 | −1 | +1 | 0 | 13.7 + 0.6 |
| 4 | +1 | +1 | 0 | 16.0 ± 0.9 |
| 5 | −1 | 0 | −1 | 13.6 ± 0.7 |
| 6 | +1 | 0 | −1 | 15.1 ± 0.7 |
| 7 | −1 | 0 | +1 | 13.4 ± 0.7 |
| 8 | +1 | 0 | +1 | 16.7 ± 0.6 |
| 9 | 0 | −1 | −1 | 12.9 ± 0.7 |
| 10 | 0 | +1 | −1 | 15.7 ± 0.7 |
| 11 | 0 | −1 | +1 | 16.6 ± 0.3 |
| 12 | 0 | +1 | +1 | 16.2 ± 0.5 |
| 13 | 0 | 0 | 0 | 18.7 ± 0.8 |
| 14 | 0 | 0 | 0 | 19.9 ± 0.9 |
| 15 | 0 | 0 | 0 | 20.1 ± 0.9 |
| 16 | 0 | 0 | 0 | 19.5 ± 0.4 |
| 17 | 0 | 0 | 0 | 20.3 ± 0.8 |
Concentrated proteolytic extract preparation
The experiment steps of crude enzyme preparation were carried out at 4 °C. The cells were harvested by centrifugation of a 2-day-old culture of S. marasensis at 8000×g for 10 min. The precipitation of the cell-free supernatant was performed by gradually adding up to an 80 % (v/v) saturation pre-cold ethanol on to the media and stirring for 2 h. The precipitate was concentrated by centrifugation at 20,000×g for 15 min and suspended in a minimal volume of 50 mM phosphate buffer (pH 8.0) including 20% NaCl. The concentrated extract was dialyzed against same buffer overnight and the proteolytic activity was measured in the assay conditions.
The biochemical and functional characterization of the proteolytic extract
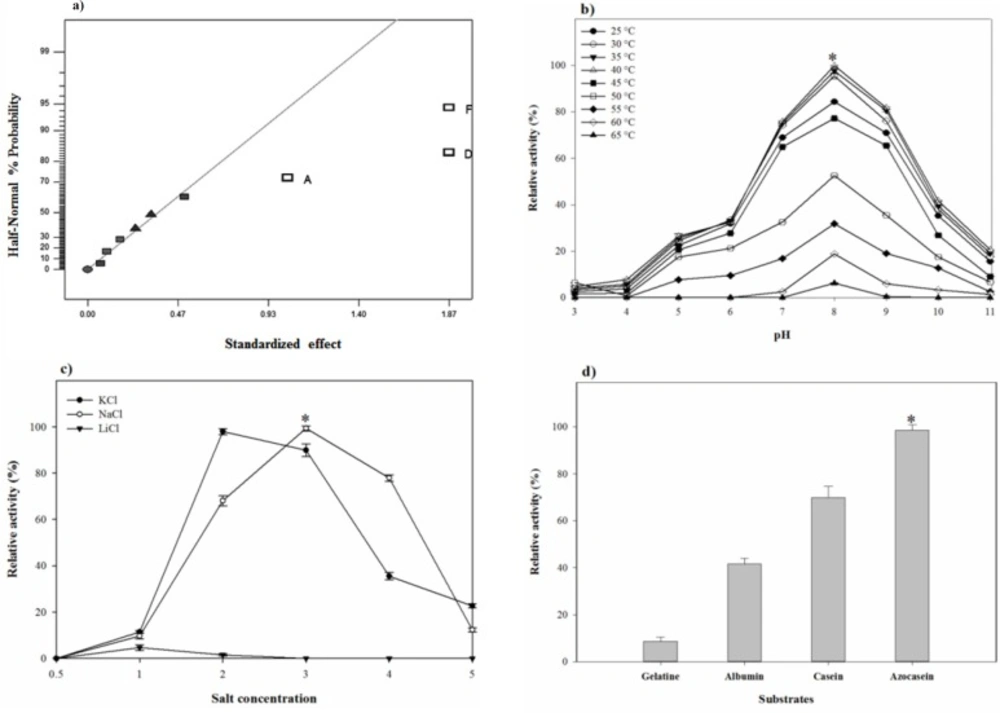
The thermal- and pH-stability of the proteolytic
The stability of the crude enzyme was investigated in a broad range of pH (3.0–11.0) and temperature (25–65 °C). The enzyme samples (~ 20 U) were adjusted to the desired pH values and were incubated at different temperatures. After an hour, the residual activity of the proteolytic extract was assayed according to the method described in the earlier section and the initial proteolytic activity before starting the pre-incubation was considered as 100%. The buffer systems for pH were citrate buffer (pH 3.0–5.5), phosphate buffer (pH 6.0–8.0), Tris-HCl buffer (pH 8.5–9.5), and glycine-NaOH buffer (pH 10.0–11.0) at 0.1 M concentration, while 20% NaCl (w/v) was incorporated into the used buffers for optimal activity.
The effects of inorganic salts and metal ions on the proteolytic stability
The enzyme sample (~ 20 U) was added to phosphate buffer (pH 8.0) containing NaCl, KCl, and LiCl with the final concentration of 0–30% in the mixture. After an hour incubating at room temperature and shaking at 150 rpm, the residual proteolytic activity assayed and the original enzyme activity existed before the incubation period was considered as 100%. The stability of the crude protease was also investigated in the presence of Fe3+, Fe2+, Ni2+, Co2+, Hg2+, Mn2+, Mg2+, Zn2+, Cu2+, Al3+, and Li+ at the concentrations of 1, 5, and 10 mM dissolved in 100 mM phosphate buffer (pH 8.0) containing 20% NaCl. The residual activity was determined after an hour of incubation at room temperature with an orbital shaking at 150 rpm. The enzyme activity in the absence of any di- and tri-valent ions was taken as 100%.
The effects of surfactants, inhibitors, and organic solvents on the proteolytic stability
The effects of several surfactants (cetyltrimethylammonium bromide (CTAB)
(1-10 mM), Triton X-100 (1-10 mM), SDS (5-20 mM), and Tween 80 (1-5 mM)), and denaturing agent of urea (at the same concentrations of CTAB and Triton X-100) in 0.1 M phosphate buffer (pH 8.0) containing 20% NaCl on the enzyme stability were also studied. In addition, the effects of protease inhibitors of PMSF (1-10 mM), EDTA (1-10 mM), and 2-ME (10-100 mM) were investigated. The mixture of protease extract (~ 20 U) and agents was incubated at room temperature with an orbital constant shaking at 150 rpm for an hour and the residual activity was assayed by the procedure. The proteolytic activity without any additives was set as 100%.
In addition, the stability of the crude protease in organic solvents with various log P values at the concentrations of 20, 50, and 80% (v/v) was studied. Organic solvents chosen in this study were acetonitrile, acetone, ethanol, butanol, hexanone, cyclohexanol, chloroform, octanol, cyclohexane, hexane, heptane, decanol, nonane, dodecane, and hexadecane. The mixture of protease extract (~ 20 U)-solvent was incubated at room temperature with a constant shaking at 150 rpm for an hour. The aqueous phase was carefully withdrawn and assayed and the residual specific proteolytic activity was applied to express the enzyme stability. Each result was compared with the specific activity of the corresponding control.
The substrate specificity analysis
The substrate specification was investigated by incubating the crude protease with several substrates including casein, azocasein, albumin, and gelatin. Each substrate was dissolved in 100 mM phosphate buffer (pH 8.0) containing 20% NaCl to a concentration of 1% w/v. Then, substrate solution was mixed with 1 mL of the supernatant containing protease (~ 20 U) to obtain a final volume solution pointed in the protease assay, previously. Indeed to test the proteolytic extract substrate specificity, the assay procedure explained above was followed except casein was replaced by the specific protein substrate and the released tyrosine was estimated.
Statistical analyses
To show the significant difference between groups, One-way ANOVA followed by Holm-Sidak multiple comparison test and two-way ANOVA were applied. Statistical significance was considered at p < 0.05.