Nonionic surfactants of PS 20 and polysorbate 80 (PS 80) are widely used in protein formulations to prevent aggregation because of their effectiveness at low concentrations, low toxicity and their ability to minimize protein surface adsorption and aggregation under various processing condition (
1-
3). Most of the marketed monoclonal antibodies formulations contain one of these surfactants (
4). PS 20 is used in some cytokines such as Actimmune1 (IFN–gamma-1b) and Avonex (IFN β_1a) as well as Ontak1 (Denileukin diftitox) (
5,
6).
Several mechanisms for protein stabilization by surfactants have been proposed. Nonionic surfactants confer surface-induced anti-aggregation behavior by competing with proteins for interfaces which could initiate aggregation (
7). Binding to hydrophobic regions on the protein surface and reduction in intermolecular interactions (
8-
10), increase in the free energy of protein unfolding (
11) and acting as chemical chaperone (
12,
13) have also been reported as possible anti-aggregation mechanisms of surfactants. They can also modulate adsorption loss and aggregation by coating interfaces and/or participating in protein–surfactant associations (
14). Inherent impurities within the surfactants such as PS 80 can also influence significantly photostability of a protein (
15). Commercial polysorbates contain a large amounts of polyoxyethylene, sorbitan polyoxyethylene, and isosorbide polyoxyethylene fatty acid esters (
16,
17,
1). These products may contain residual amounts of peroxides that in aqueous solution could react and damage proteins (
18-
20).
Therapeutic proteins are prone to be oxidized during different manufacturing steps and storage, due to exposure to intense light, trace amounts of metal ions or peroxide impurities in polysorbate excipients (
21-
23). Amino acids containing either sulfur atom (cysteine and methionine) or an aromatic ring (histidine, tryptophan, tyrosine and phenyl alanine) are at most risk of oxidation (
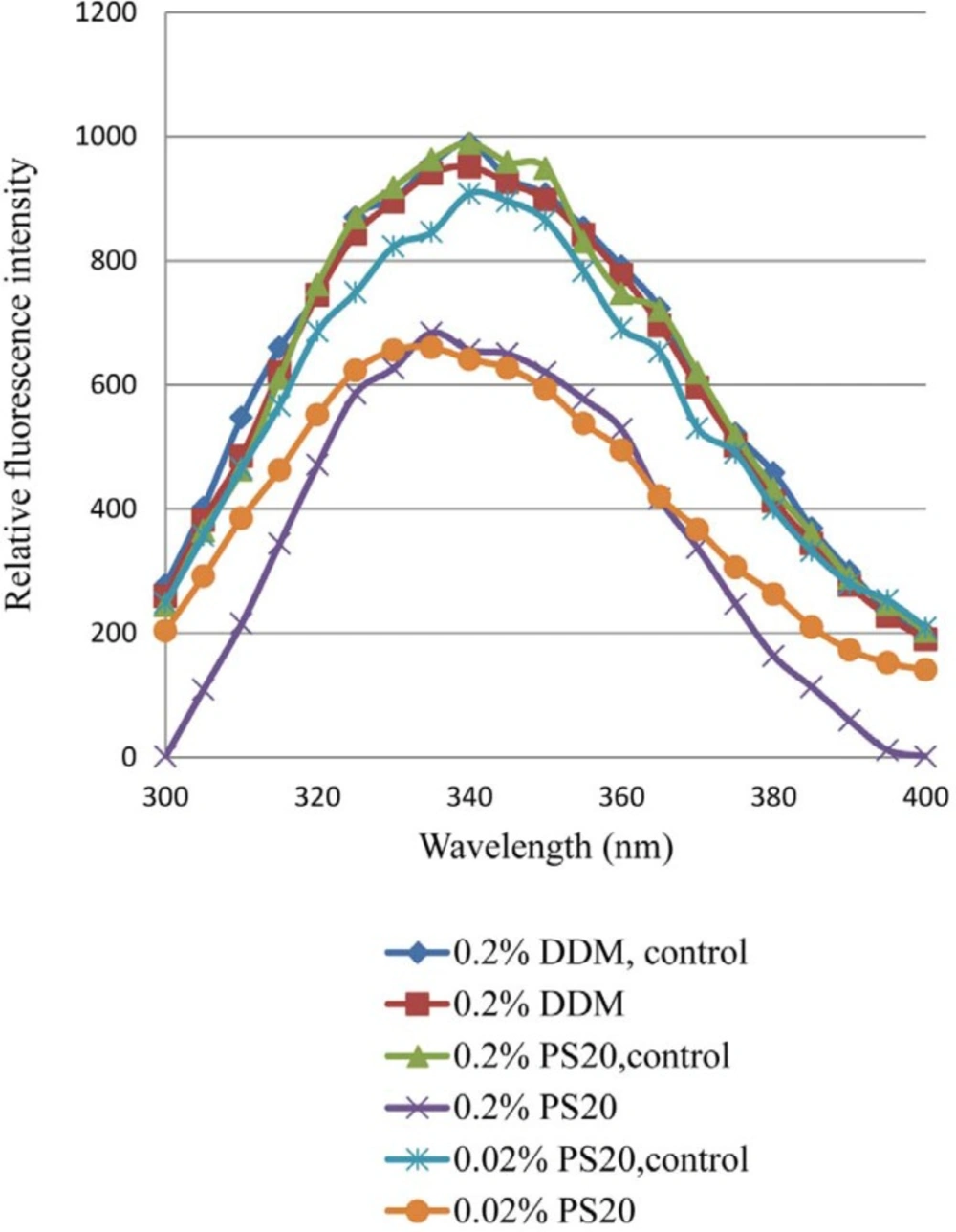
24). For the analysis of protein circular dichroism, it is observed amino acids with aromatic functionality in protein such as tryptophan (Trp) is likely to make significant contribution. With a light source at an emission spectrum of greater than approximately 300 nm, Trp is the major amino acid residue in proteins with significant absorbance (
25,
26). More often oxidation increase the propensity to aggregates. Oxidation of Trp residues and subsequent aggregation of the model therapeutic protein type I soluble tumor necrosis factor receptor has been reported to be induced by UV light at 302 nm (
5). Peroxodisulfate-mediated oxidation of recombinant human interleukin-2 altered secondary structure of the protein, as observed by far-UV CD (
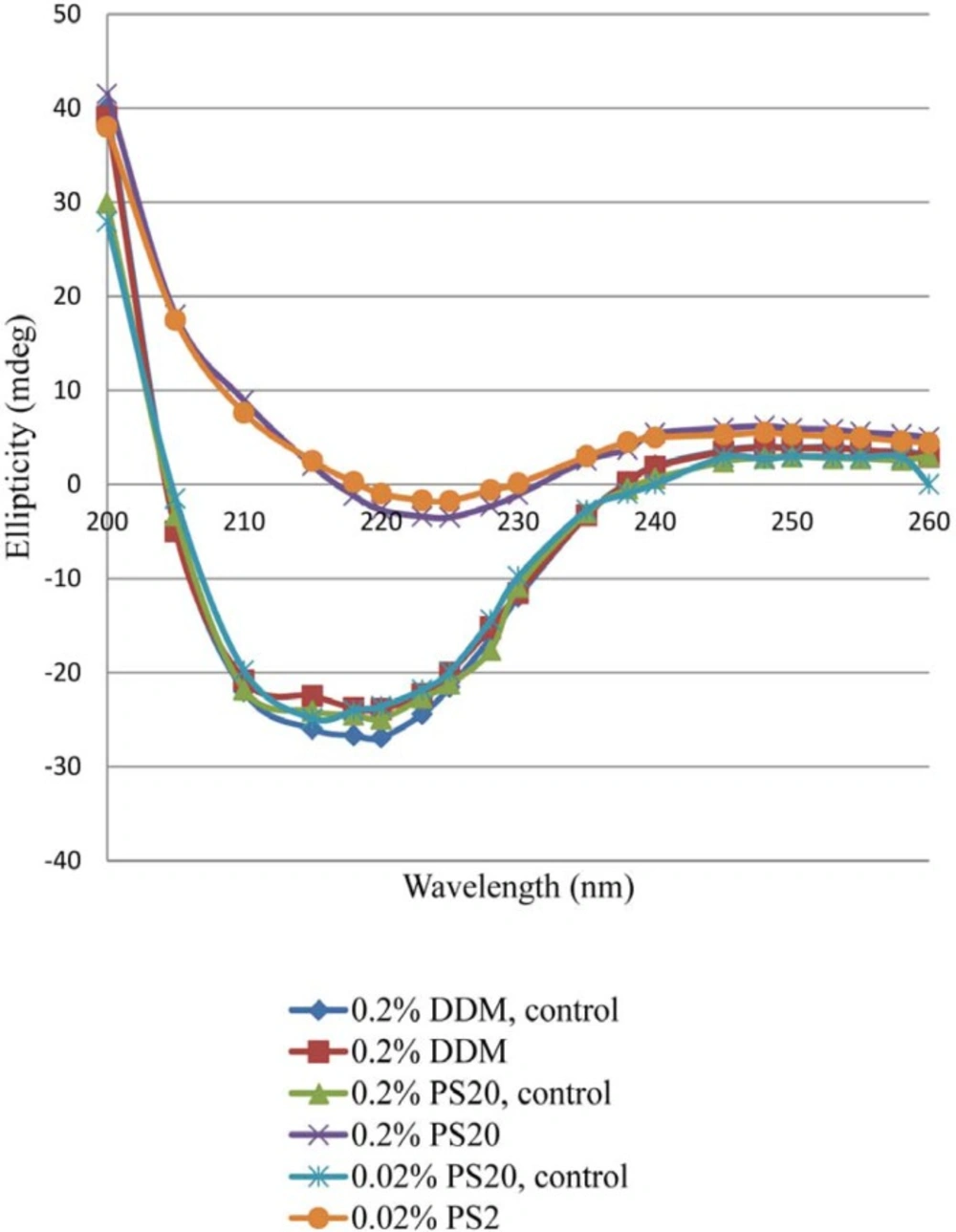
27).
Potential problems with using polysorbates and other polyoxyethylene-containing surfactants make clear the need for alternative surfactants that prevent aggregation and unintended protein damage. To circumvent the effect of residual inherent oxidative species within the surfactants, one of alternatives is a class of non–ether-based alkylsaccharide surfactants composed of a sugar moiety coupled to an alkyl chain that show significant improvement in stability and reduced immunogenicity (
28). A broad range of alkylsaccharides (nonionic surfactants) comprise_Intravail alkylsaccharide excipients, have been studied as transmucosal absorption facilitator (
29-
31). It was observed that some of these molecules such as DDM have significant anti-aggregation effect (
28). n-Dodecyl-β-D-maltoside is a non-toxic, non-mutagenic, and non-irritating and nonionic suger-based surfactant (
28,
32) that finally metabolizes into CO2 and H2O through the corresponding sugar and fatty acid (
28,
32).
Although there are several liquid protein formulations in the market that have used PS 20 as an anti-aggregation agent (
6,
33), incorporation of DDM in biopharmaceutical formulation has not yet received much attention due to scarce research.
However, one problematic issue in using polysorbates for biopharmaceutical is their potential adverse effect on protein stability, which has not been extensively reported. There are few examples of oxidate damage of recombinant human ciliary neurotrophic factor in solution and recombinant human granulocyte colony-stimulating factor in solution due to presence of residual peroxides in PS 80 (
34). Specific products have recently been addressed, including monoclonal antibodies, calcitonin, granulocyte colony-stimulating factor, growth hormone, insulin, interferon alpha and beta, oxytocin and parathyroid hormone (
35). The importance of different concentrations of PS 80 as a polyoxyethene-containing additive in formulation of Monoclonal antibody MAb1 has been investigated under stress condition (
36).
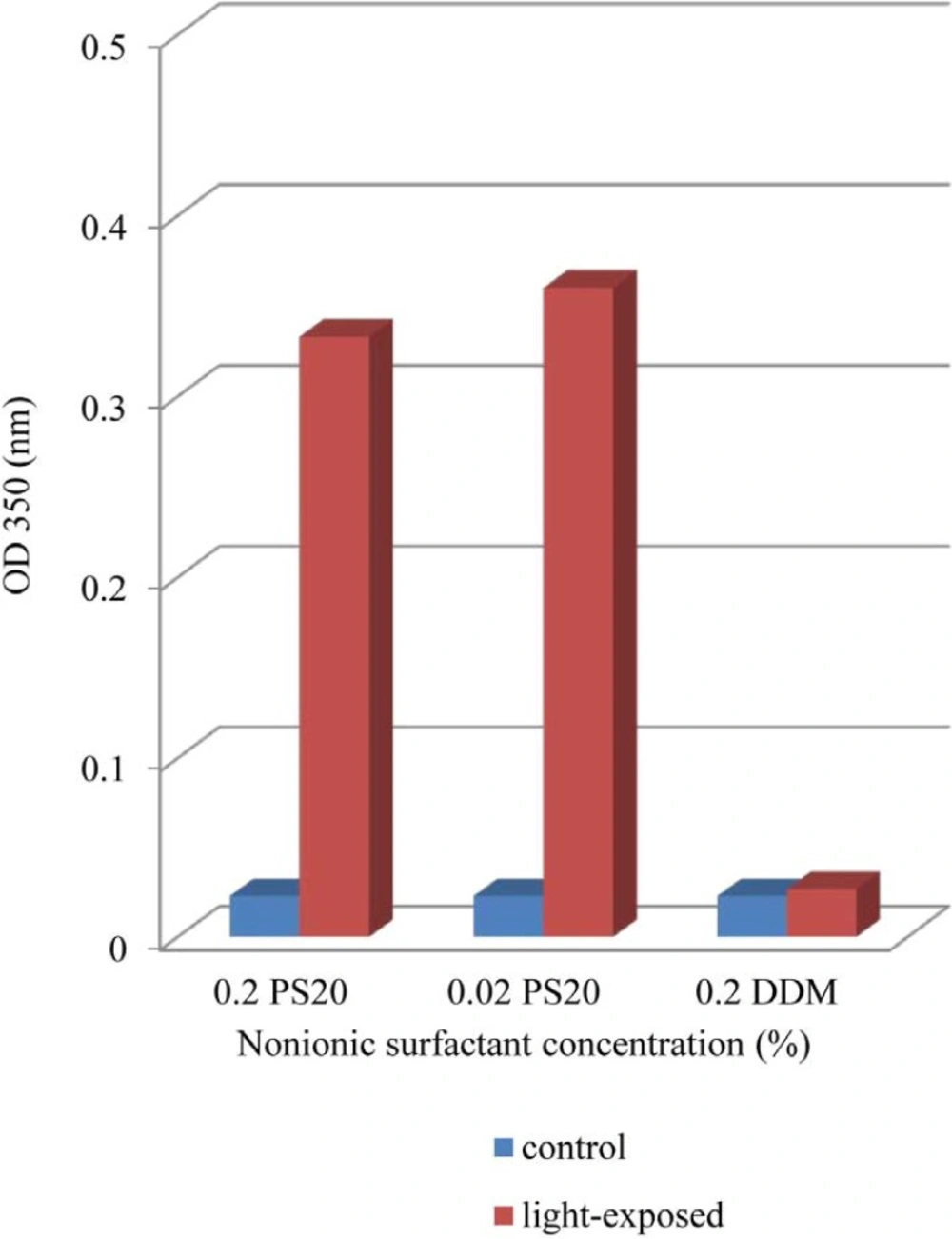
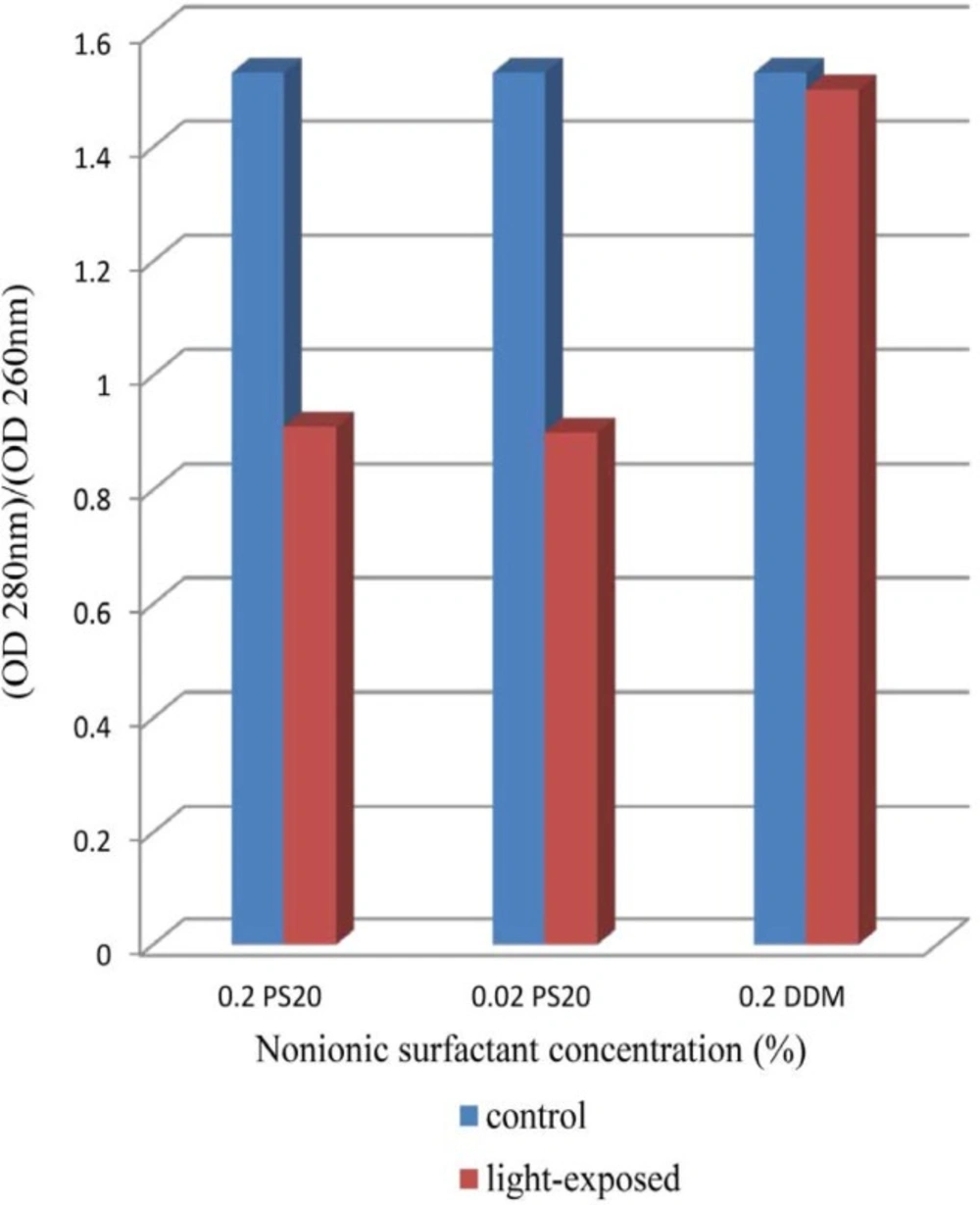
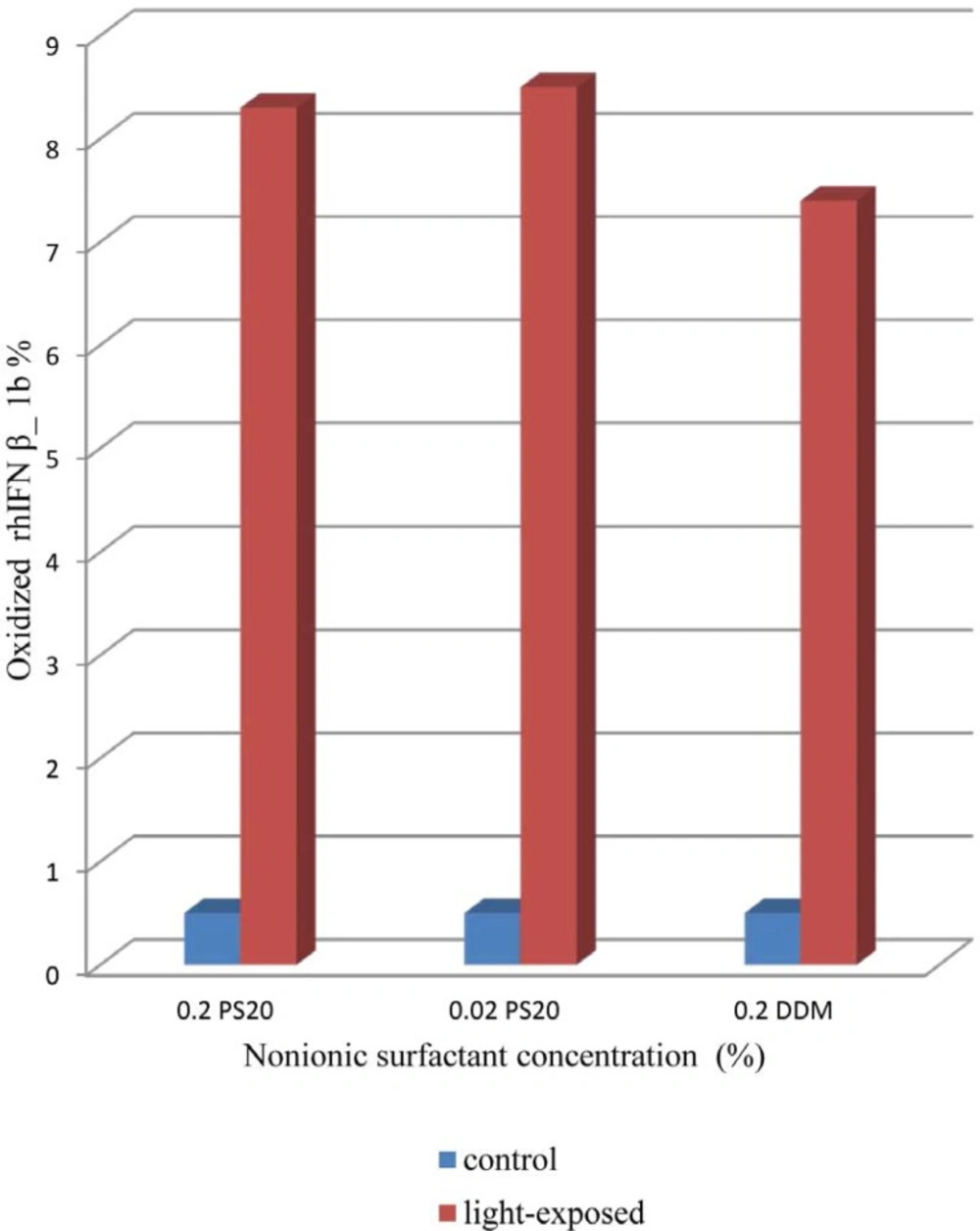
Effect of non-ionic surfactants (DDM and PS 20) on prevention of aggregation and conformational changes in liquid formulation of recombinant human IFNβ_1b induced by light has been explored in current study.