With advances in cancer biology, the cancer stem cells or tumor-initiating cells were identified as pluripotent cells with a unique capacity of self-renewal being able to differentiate into a heterogeneous population of cells to form tumors. Technical progress over decades led to purification and identification of CSCs based on specific surface markers. Biomarkers such as CD44, CD24, Cytokeratin 5 (CK5), and SOX4 have been proposed for isolation and characterization of breast CSCs (
27). It has been shown that breast CSCs are multiple, distinct, and non-overlapping populations co-existing within the tumor mass. Since a minute fraction of tumor cells includes CSCs, their characterization usually mediated through isolation using cell surface markers and development
in-vitro (
28). Most current studies have demonstrated CD44 and CD24 as proposed markers for isolation of CSC subset in breast tumors (
29). In our previous study, we isolated CD44
+/CD24
-/low breast CSCs from the main population of MDA-MB-231 cell line using MACS and evaluated the CD44/CD24 expressions by flow cytometry. After isolation, the percentage of subpopulation expressed CD44
+/CD24
-/low biomarkers elevated significantly from 51.10% to 82.24% (
26). In addition, it was reported that isolated CSCs exhibit more tumorigenicity
in-vivo and resistance to conventional chemotherapy and radiotherapy than their parental cells. Therefore, cancer stem cell-based therapies are being investigated as a promising avenue to successful cancer treatment (
28).
Recently, investigations clarified that cannabinoids inhibit cell proliferation, induce apoptosis and harness cell migration and angiogenesis in various cancer cells (
30). Vara and colleagues found that treatment of hepatocellular carcinoma cells with THC which is the active component of
Cannabis sativa decreases cell viability and tumor growth via autophagy
in-vitro and
in-vivo (
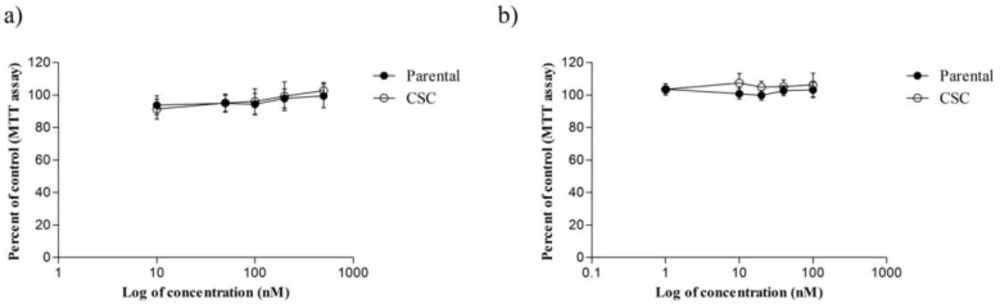
31). However, our MTT assay showed that ACEA and AM251 do not have an anti-proliferative effect on CSCs and MDA-MB-231. Thus, no correlation between the anti-invasion effect of cannabinoid agonist ACEA and cytotoxic effects of the drug was found and the specific cannabinoid receptors may be involved (
32).
Analyzing the relation between ligand doses in the target zone and biological response obtained are among the most remarkable research fields with many clinical applications. Unlike previous studies on cannabinoids applied AM251 at micromolar doses (
33), the present study used nanomolar doses of AM251. Studies indicated that nanomolar doses of cannabinoids may function as growth factor-like molecules in autocrine and/or paracrine manner and accelerate tumor growth (
34) while cannabinoid agonists at micromolar concentration reduce tumor cell proliferation in a dose-dependent manner (
35).
The results of MTT assay on human MDA-MB-231 cancer cells in the presence of several cannabinoid antagonists illustrated that these compounds increase cell proliferation at nanomolar concentration. Besides, at 10 µM of methanandamide (an agonist of CB receptors and vanilloid receptor 1), cell proliferation significantly increases. These studies showed the proliferative effects of cannabinoids at low doses (
32).
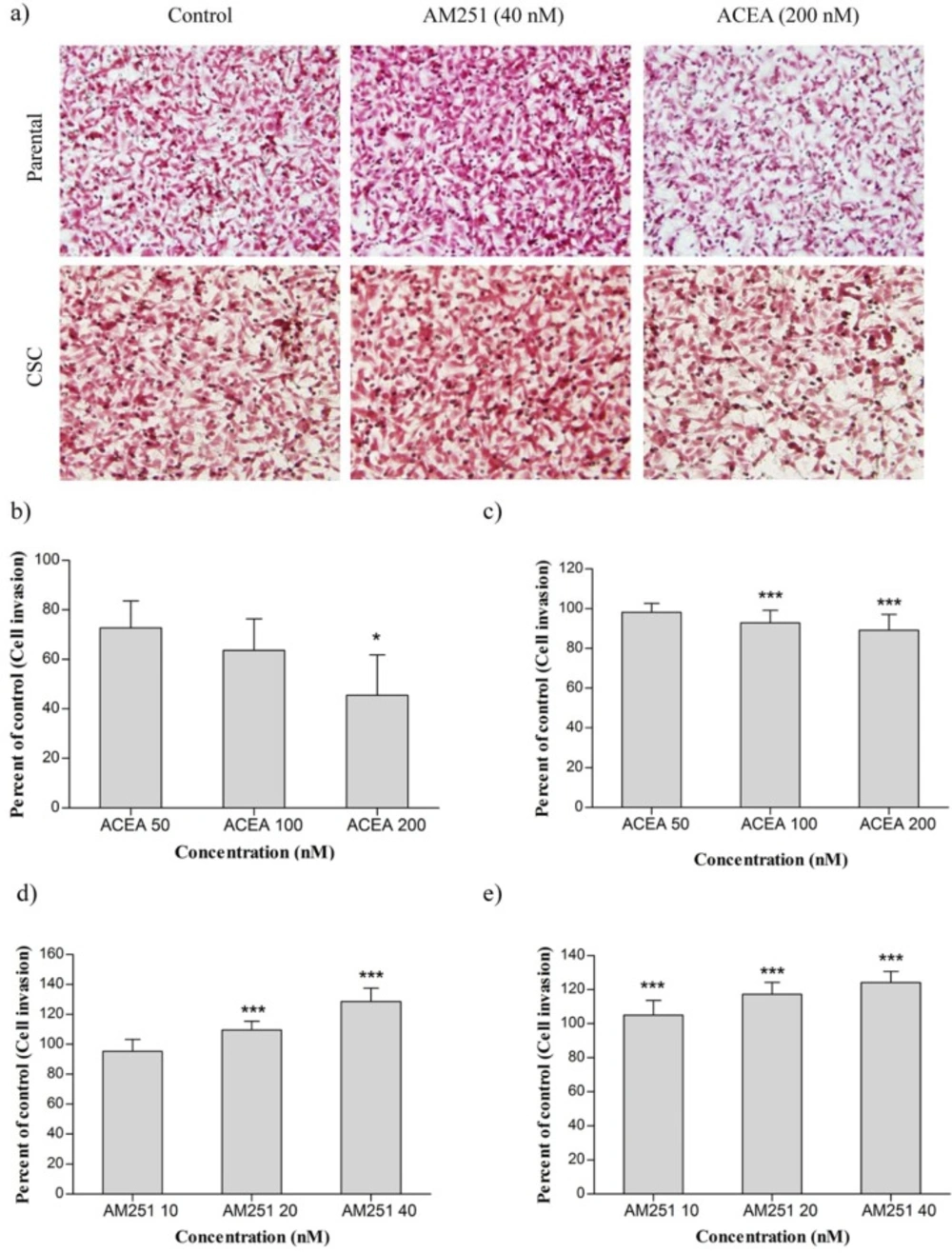
Extensive studies have reported that Epithelial-to-Mesenchymal Transition (EMT) promotes invasion and migration of carcinoma cells. Induction of EMT features lead to cell reprogramming and generate cells with CSC properties and invasive potential required for metastatic spread (
25). Our data showed that ACEA has an inhibitory effect on CD44
+/CD24
-breast cancer stem cell and their parental cell invasion but AM251 increases invasiveness. It is important to mention that CD44, a transmembrane glycoprotein, can interact with hyaluronic acid, fibronectin, fibrinogen, collagen, laminin, fibroblast growth factor-2, osteopontin, and MMPs which is correlated with cell migration (
36). Blázquez
et al. demonstrated that THC administration interferes with MMP2 expression and prevents cell invasion in glioma mouse model (
37). In addition, treatment of MDA-MB-231 cells with ACEA inhibits MMP2, VEGF, and cyclooxygenase-2 expression via CB1 receptors which have expressed in MDA-MB-231 cells as confirmed by RT-PCR and western blot (
5,
32). Of special note, CB1 cannabinoid receptor is responsible for anti-angiogenic effects and inhibition of cell invasion in human metastatic MDA-MB-231 breast carcinoma cells (
32).
In general, cannabinoid agonists lead to CB1 receptor overexpression however a similar response has been observed in various cancer types (
18). Studies demonstrated that increased cannabinoid receptor expression happens upon THC treatment in cancer cells compared to normal cells. The mechanism of this phenomenon has not elucidated yet but it has been recognized that there is an important correlation between cannabinoid receptors and cancer (
38). The existence of CB1a and CB1b (splice variants of CB1) could reflect the different response of cannabinoid receptors in malignant and normal cells (
39).
In summary, our results clarified that cannabinoid receptor agonist possesses anti-invasion potential in both main population and breast cancer stem cells while AM251 exhibits converse effects. Additionally, ACEA and AM251 did not show any cytotoxicity towards CSCs and MDA-MB-231 parental cells. Considering that most anti-cancer drugs do not eradicate stem cells and only target main population cells, the results disclosed here can be used for prevention of cancer recurrence.