
Introduction
Experimental
Results
Discussion
| Healthy Subjects | CAD Patients | p-value | |
|---|---|---|---|
| Characteristics | Median ± IQR | Median ± IQR | Mann-Whitney test |
| Age (years) | 50 ± 2 | 51 ± 4 | 0.12 |
| Body Mass Index (Kg/m2) | 25.1 ± 2.20 | 24.95 ± 1.80 | 0.249 |
| Waist Circumference (cm) | 78.5 ± 9 | 80.5 ± 13 | 0.368 |
| Total Cholesterol (mg/dL) | 173 ± 7 | 188 ± 17 | 0.016 |
| HDL Cholesterol (mg/dL) | 40.5 ± 16 | 33 ± 16 | 0.005 |
| LDL Cholesterol (mg/dL) | 92 ± 5 | 98 ± 18 | 0.04 |
| Triglyceride (mg/dL) | 136 ± 31 | 166 ± 44 | 0.002 |
| Systolic Blood Pressure (mmHg) | 120 ± 5 | 140 ± 15 | <0.001 |
| Diastolic Blood Pressure (mmHg) | 80 ± 0 | 90 ± 5 | <0.001 |
| Fasting Blood Sugar (mg/dL) | 88.5 ± 12 | 93.5 ± 4.40 | 0.03 |
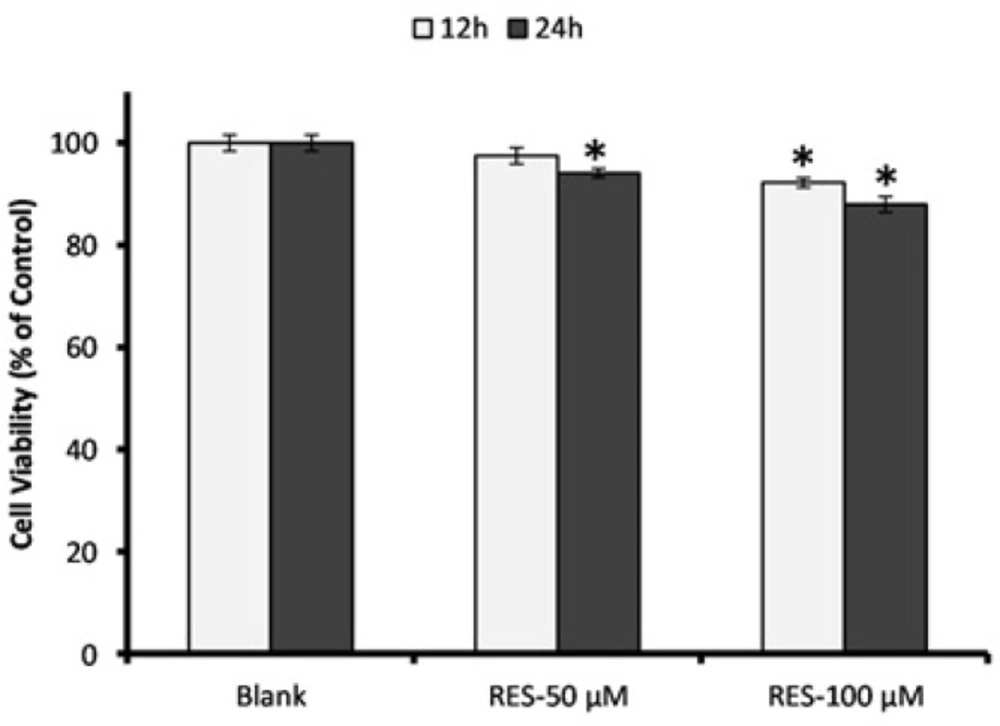
Assessment of PBMCs viability in response to resveratrol (RES) treatment. Cells were incubated with 50 and 100 µM RES for 12 h and 24 h, and cell viability was determined by MTT assay. All RES treatments, except RES 100 µM for 24 h, did not reduce the cell viability less than 90%. The MTT assay was performed in triplicate and data are reported from the mean of triplicates. Data are expressed as means ± SEM. * stands for p < 0.01.
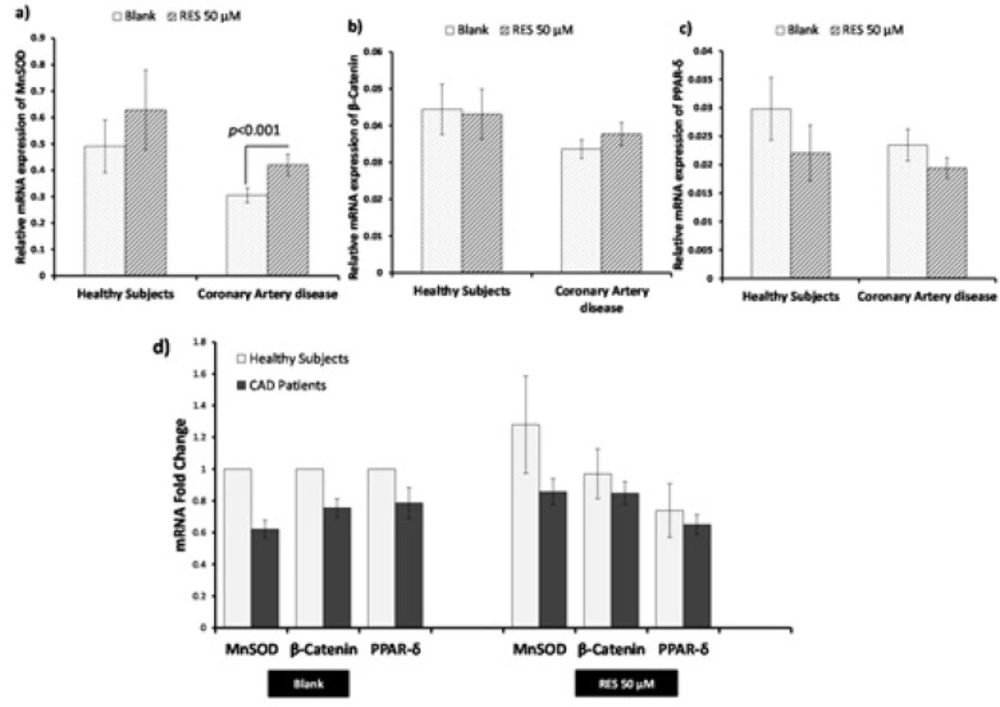
Effects of resveratrol (RES) on β-catenin, MnSOD, and PPAR-δ gene expression by real-time PCR. PBMCs were treated with 50 µM RES for 12 h incubation. a) A non-significant change in MnSOD mRNA was observed in healthy subjects after treatment with RES as compared to blank. Treatment with RES could significantly increase the mRNA level of MnSOD in CAD patients compared to blank. b) No significant differences were observed for β-catenin mRNA in healthy subjects and CAD patients after treatment with RES compared to their blank. c) No significant differences in PPAR-δ mRNA were observed for both healthy subjects and CAD patients after treatment with RES compared to their blanks. d) Between-group analysis showed non-significant higher levels of MnSOD, β-catenin and PPAR-δ in healthy subjects than CAD patients in both blank and RES treatment. RES was dissolved in DMSO, and blank groups (untreated cells) were containing only DMSO. In both blank and RES treated cells, DMSO was present at equal concentration (0.025%). All gene expression tests were performed in triplicate in each experiment, and the mean of duplicates were used for statistical analyses. Data are expressed as means ± SEM.
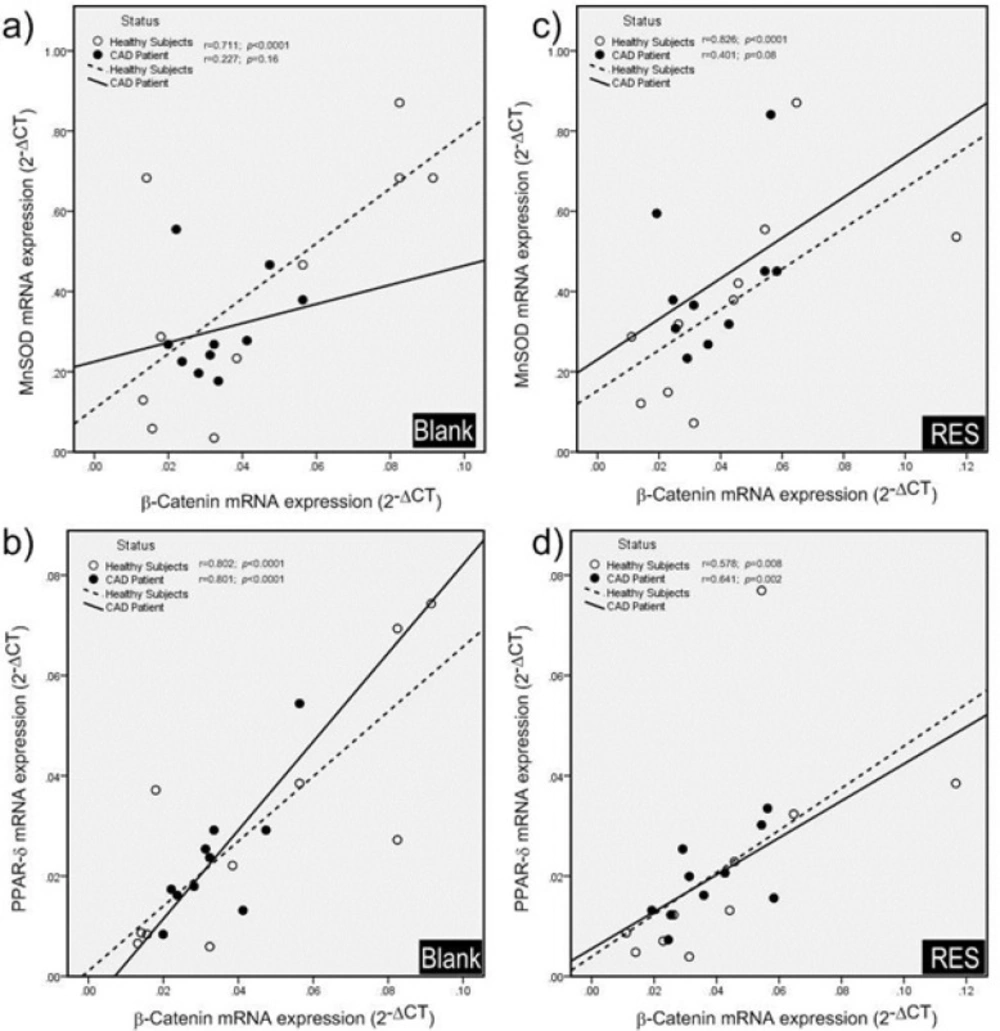
Pearson correlation coefficients of mRNA expression of genes (β-catenin, MnSOD and PPAR-δ). a) In blank groups; β-catenin and MnSOD mRNA expressions were significantly correlated in healthy subjects, while they were not significantly correlated in CAD patients. b) β-catenin and PPAR-δ mRNA showed significant positive correlations in both healthy subjects and CAD patients in blank groups. c) After treatment with resveratrol (RES) a significant positive correlation was found between β-catenin and MnSOD mRNA expressions in healthy subjects, but it was not observed in CAD patients. d) Also β-catenin and PPAR-δ mRNA showed significant positive correlations in both healthy subjects and CAD patients after treatment with RES. RES was dissolved in DMSO, and blank groups (untreated cells) were containing only DMSO. In both blank and RES treated cells, DMSO was present at equal concentration (0.025%).
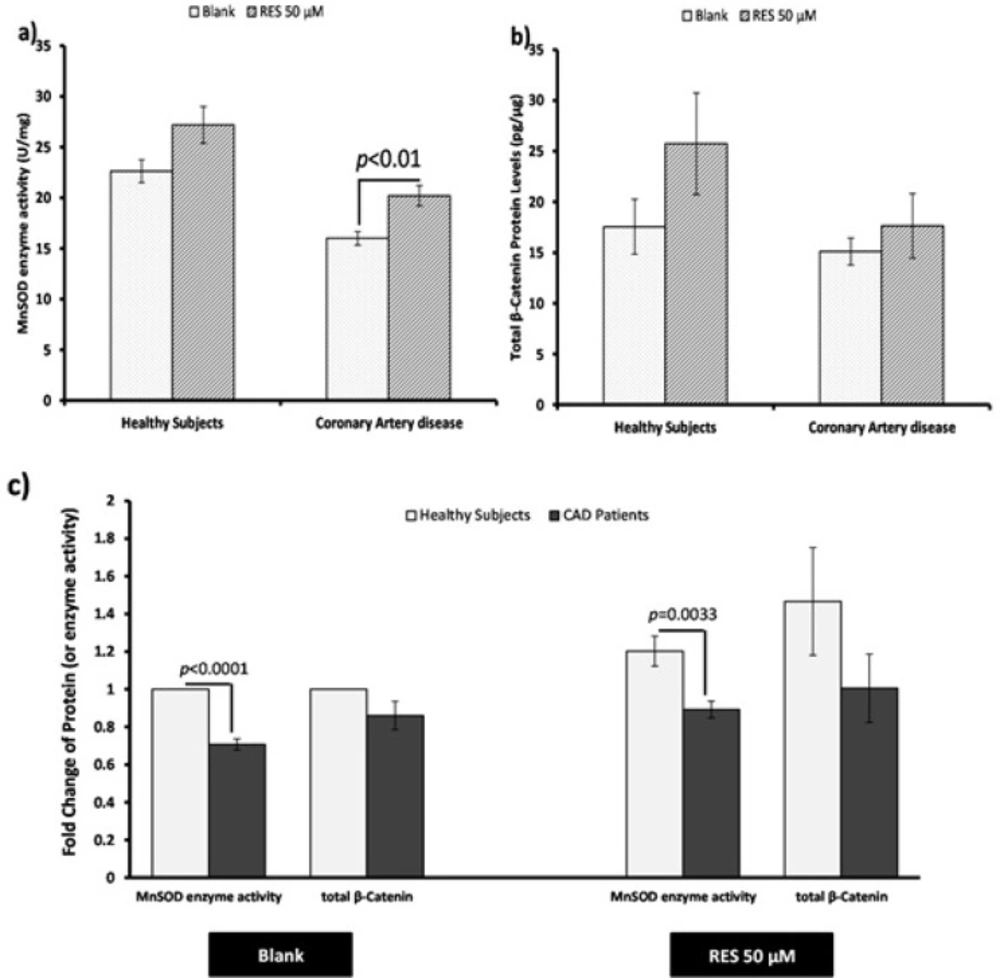
Effects of resveratrol (RES) on MnSOD enzyme activity and total β-catenin protein. PBMCs were treated with 50 µM RES for 12 h incubation. a) MnSOD enzyme activity was non-significantly increased in healthy subjects after treatment with RES. Also it was significantly increased in CAD patients after treatment with RES in comparison with blank. b) After treatment with RES a non-significant increasing trend was found for total β-catenin protein of healthy subjects compared to blank. Total β-catenin protein of CAD patients did not significantly increase after treatment with RES. c) Between-group differences showed higher MnSOD enzyme activitiy and total β-catenin protein levels for healthy subjects in comparison with CAD patients. RES was dissolved in DMSO, and blank groups (untreated cells) were containing only DMSO. In both blank and RES treated cells, DMSO was present at equal concentration (0.025%). All experiments were performed in duplicate, and the mean of duplicates were used for statistical analyses. Data are expressed as means ± SEM
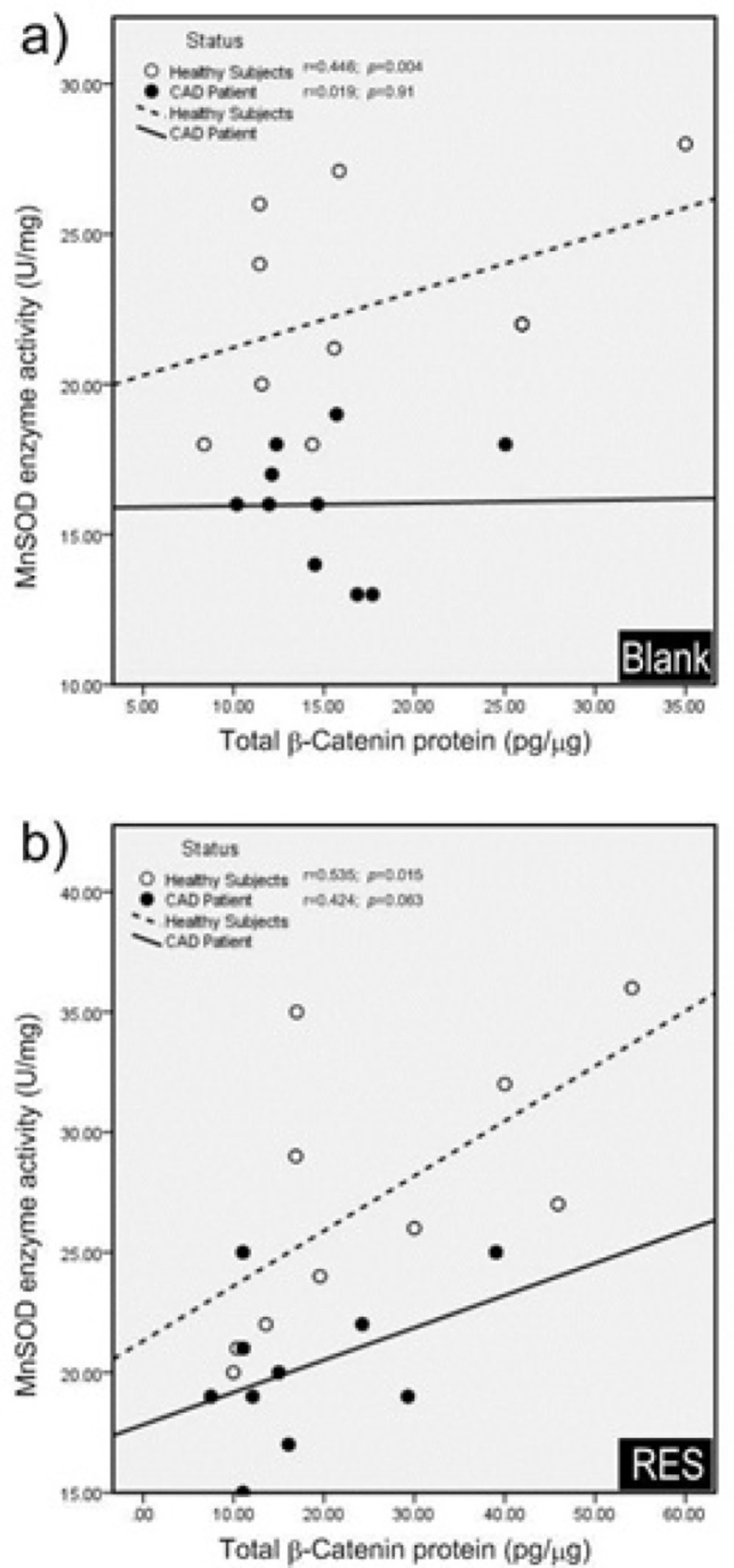
Pearson correlation coefficients of total β-catenin protein and MnSOD enzyme activity. a) In blank group; total β-catenin protein and MnSOD enzyme activity were significantly correlated in healthy subjects, while they were not significantly correlated in CAD patients. b) After treatment with resveratrol (RES), total β-catenin protein and MnSOD enzyme activity showed a significant positive correlation in healthy subjects, while no significant correlation was observed in CAD patients. RES was dissolved in DMSO, and blank groups (untreated cells) were containing only DMSO. In both blank and RES treated cells, DMSO was present at equal concentration (0.025%)
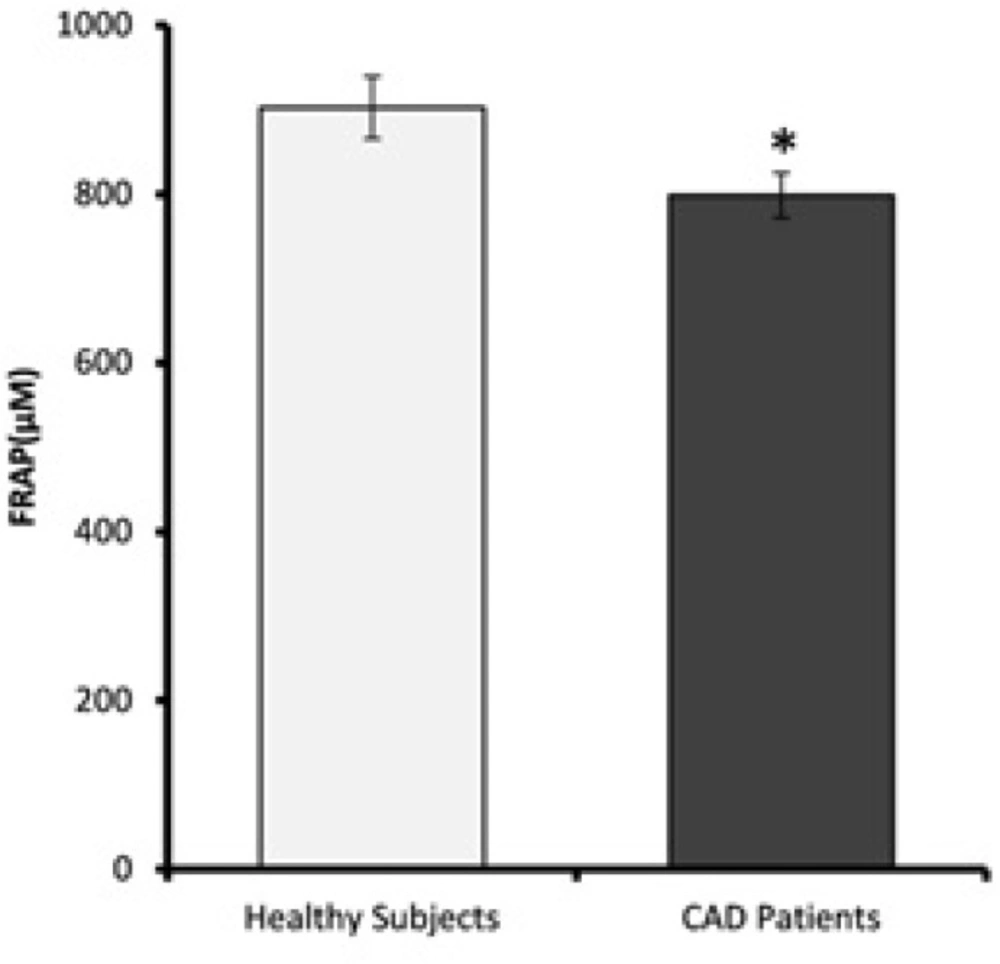
Plasma ferric reducing antioxidant power (FRAP) values. FRAP levels of CAD patients are significantly lower than healthy subjects. Data are expressed as means ± SEM. * p = 0.02
Effects of resveratrol (RES) on β-Catenin/Wnt and FOXO signaling pathways in PBMCs of CAD patients. The disrupted β-Catenin/FOXO pathway in CAD patients might be due to prolonged exposure to oxidative stress, which exerts a pathogenic role in CAD. RES could increase the MnSOD enzyme activity, however it was not through the β-Catenin/FOXO pathway. The β-catenin/Wnt pathway was intact in CAD patients but was not provoked by RES. (LRP5/6, Low density lipoprotein receptor-related protein 5/6; APC, Adenomatosis polyposis coli; GSK-3β, glycogen synthase kinase-3β; CK1, Casein kinase 1; TCF/LEF, T-cell factor/lymphoid enhancer factor; FOXO, Forkhead box O; MnSOD, Manganese superoxide dismutase; PPAR-δ, Proxisome prolifrator-activated receptor delta
