Bacterial strains and culture conditions
Eleven strains of
S. aureus were used for this study (
Table 1). T1 (FRI1189S) was originally isolated from a case of menstrual toxic shock syndrome (MTSS) and supplied by M. S. Bergdoll; T4 was from a burned patient with confirmed non-menstrual toxic shock syndrome (NMTSS, Bury and Rochdale Healthcare Trust, UK), RN4282 was obtained from T.J. Foster, University of Dublin, Ireland and 8325-4 from R. Novick, New York University of Medicine, USA. All other strains were isolated from burn wounds (
12).
S. aureus RN4282, T4, B6, B23, B45 and B54 were responders, T1, B3 and B35 were non-responders, 8325-4 had not been tested and B10 did not produce TSST-1 and was included as a control. All strains were confirmed as
S. aureus by performing Gram stain, catalase, oxidase and Staphaurex (Murex Biotech Limited, Kent, UK) tests.
| Strain | Gender/Age | IsolationSite of Burn | Toxin | Sapi | Responder Status |
|---|
| T1 | unknown | MTSS | TSST-1 | SAPI2 | NR |
| T4 | unknown | NMTSS | TSST-1 | SAPI2 | R |
| B3 | M/3y | Right arm | TSST-1 | SAPI2 | R |
| B6 | M/4y | Hand | TSST-1 | SAPI2 | NR |
| B10 | F/10m | Chest | Ent C | | NT |
| B23 | M/unknown | Chest | TSST-1/ Ent A | SAPI2 | R |
| B35 | M/11y | Skin | TSST-1 | SAPI2 | NR |
| B45 | M/11y | Abdomen | TSST-1 | SAPI1 | R |
| B54 | F/unknown | unknown | TSST-1 | SAPI1 | R |
| 8325-4RN4282 | Laboratory strainLaboratory strain | TSST-1TSST-1 | SAPI1 | NTR |
Kinetics of exoprotein production in sub-inhibitory concentrations of silver sulphadiazine
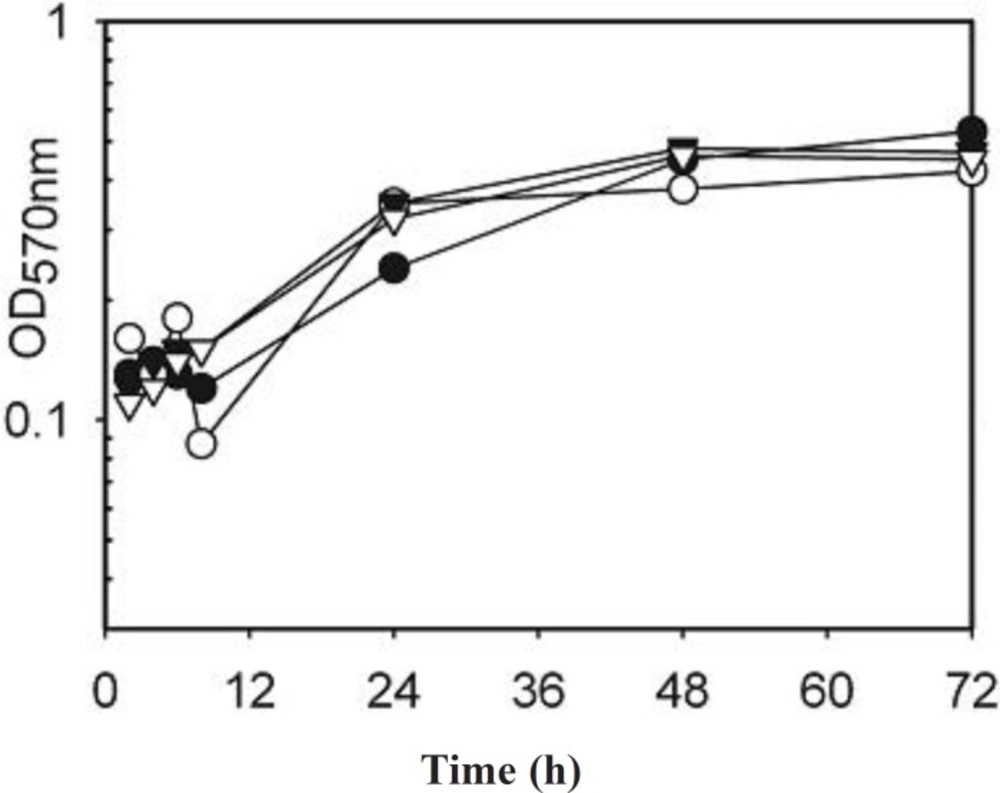
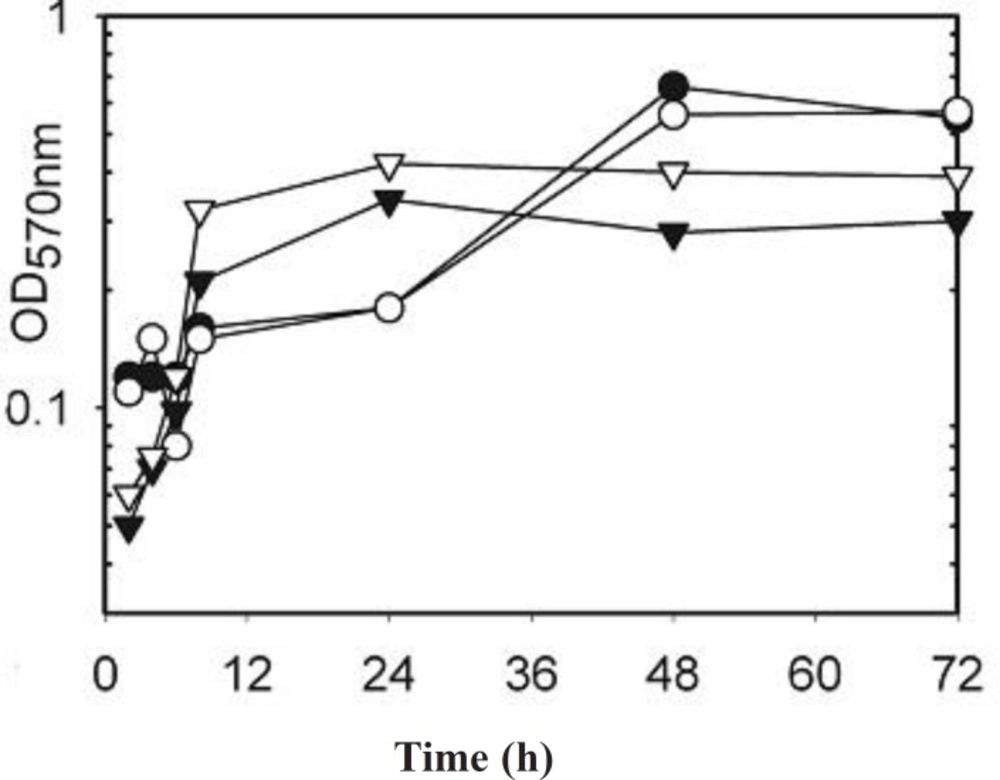
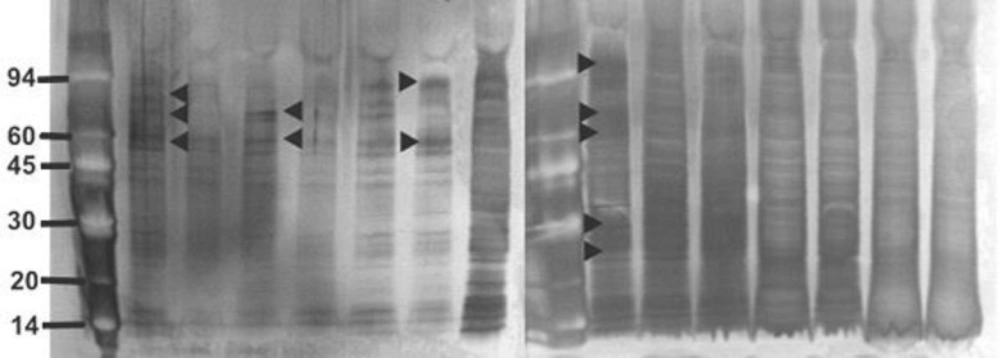
Overnight broth cultures of S. aureusμ-1 of AgSD and incubated at 37 °C as before. Flasks were removed at 2, 4, 8, 24, 48 and 72 h, turbidities were determined using a Unicam SP600 spectrophotometer at 550 nm using distilled water as a blank. Cultures with optical densities above 0.7 were diluted 1:10 with distilled water. The remainder was centrifuged for 20 min at 1500 x g and 4 °C and the supernatant fluids removed and stored at –20 °C.
Preparation of protein
Ice-cold propanol-2 (100 mL) was added to 100 mL of culture supernatant fluid in a 250 mL centrifuge tube, kept on ice for 30 min, and centrifuged at 10,000 rpm for 15 min. The pellet was collected and washed with 50% (v/v) of propanol-2 in electrophoresis buffer and centrifuged again at 10,000 rpm for 15min.The pellet was collected and dissolved in 10 mL of electrophoresis buffer (EB; 10 mM Tris-HCl, 1mM EDTA, both Sigma-Aldrich, UK, pH 8). The sample was placed in a dialysis tube (Medical International Ltd, UK) and dialysed against EB for at least 2 h at 5 ºC. The preparations were removed and stored at 0 ºC until used for electrophoresis.
Electrophoresis
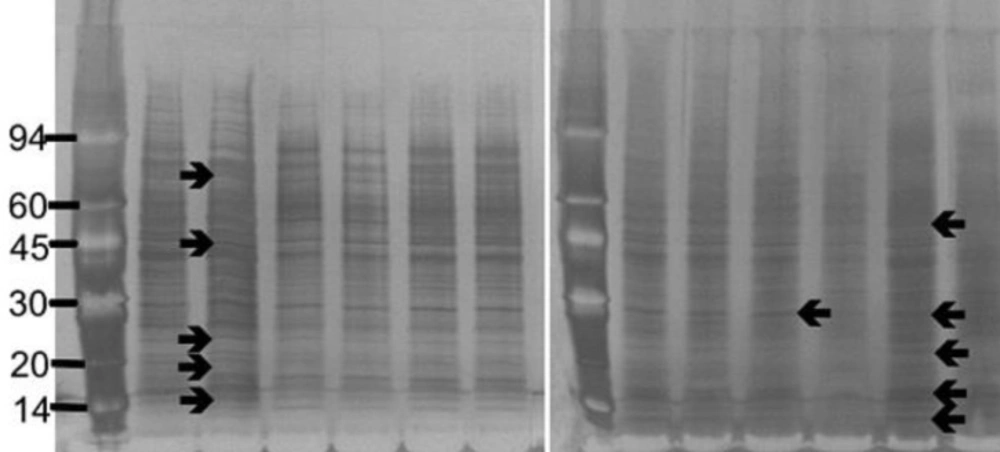
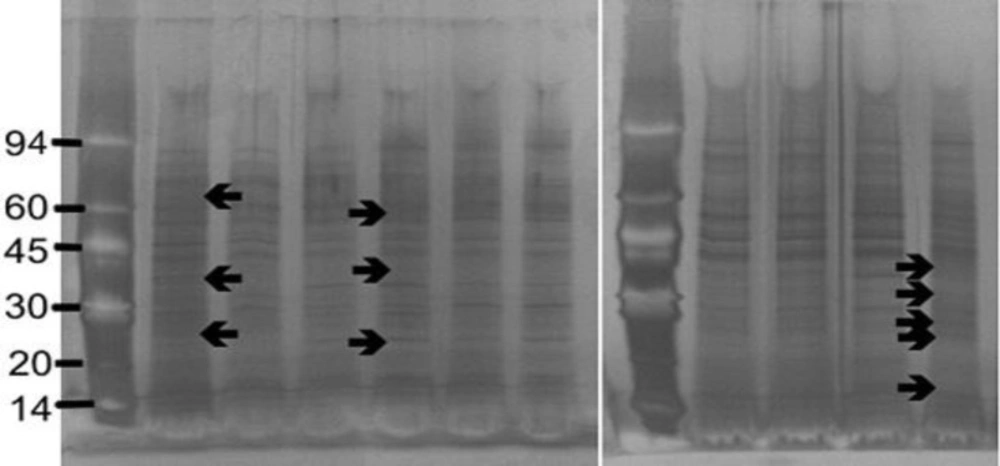
Sample (250 μL) was mixed with 250 μL of 5 % (w/v) SDS and 10 % (v/v) β-mercaptoethanol in electrophoresis buffer solution (Pharmacia Biotech, UK) in a 1.5 mL Eppendorf tube and boiled for 5 min. SDS-PAGE (Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis) was performed using Phastgel 10-15 gradient polyacrylamide gels (Pharmacia) with a continuous 10-15 % gradient and 2 % cross-linking. Protein preparations (3 µL) were applied and electrophoresis performed at 250V (10.0 mA) for 60 Vh. Protein markers (α-lactalbumin 14,000; trypsin inhibitor, 20,000; carbonic anhydrase, 30,000; ovalbumin, 43,000; bovine serum albumin, 67,000; phosphorylase b, 94,000: Amersham Biosciences, UK) were included on each gel for calibration.
Detection of proteins
The proteins were detected by staining with silver stain (Amersham Biosciences, UK). All staining solutions were prepared freshly before use as direction provided with the kit. The gels were washed x 3 with distilled water for two min on a rotary shaker. Dry gel solution (35 mL; Invitrogen Ltd) was added and shaken for 15 min. The gel was placed between two cellophane sheets (Invitrogen Ltd) and left overnight to dry.
The gels were photographed using an Alpha Imager™2200 system (Alpha Innotech Corporation, UK) and the Rƒ values and the molecular weight of each protein band was automatically calculated using the Alpha Imager™2200 and Alpha Ease™ software using the protein markers to calibrate each gel.
Estimation of protein concentration
Protein concentration was measured by the Bradford method (
14) using Bradford Reagent and Bovine serum albumin as standard (both from Sigma Aldrich, UK).