
Introduction
Experimental
Results
| Group | n | Body weight(g) | Liver weight(g) | Liver(g)/Body(g) weight×100) |
|---|---|---|---|---|
| control | 12 | 393.41 ± 24.63 | 12.36 ± 1.26 | 3.21 ± 0.30 |
| model | 12 | 272.50 ± 14.26 | 21.19 ± 2.37 | 7.78 ± 0.81 |
| group-L | 12 | 302.75 ± 18.87* | 18.11 ± 2.07* | 6.01 ± 0.80* |
| group-M | 12 | 319.17 ± 29.49* | 16.30 ± 1.73* | 5.16 ± 0.83* |
| group-H | 12 | 324.67 ± 33.96* | 15.25 ± 1.52* | 4.75 ± 0.73* |
p < 0.01 compared with group model.
p < 0.01 compared with group model.
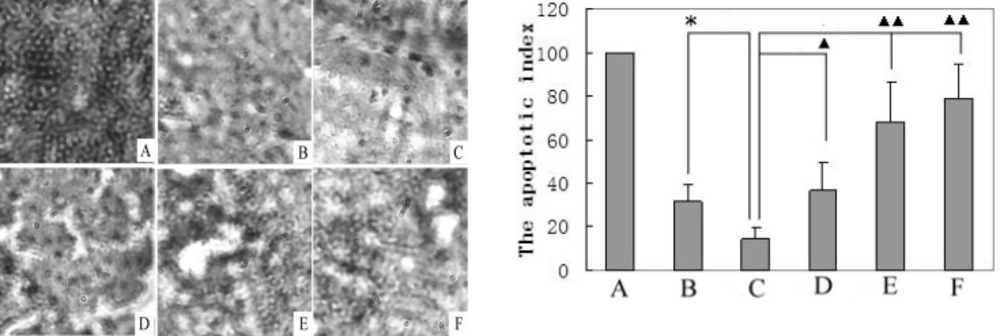
A TUNEL analysis of cell apoptosis (400×) was performed in the liver tissues of control, model, and matrine-treated rats, and their apoptotic indexes were calculated. A. Positive control (slice obtained from the apoptosis kit, set to 100%); B. Normal control group; C. Model control group; D. Matrine-treated group-L; E. Matrine-treated group-M; and F. Matrine-treated group-H. *P < 0.05 vs the control group; ▲P < 0.05 and ▲▲P < 0.01 vs the model group
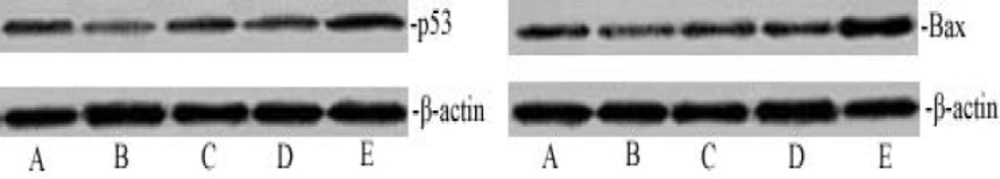
Activation of cleaved capase-3 expression depends on the p53 and Bax pathway. The dose-dependent effect of matrine on p53 and Bax expression. The cell lysates were analyzed by western blotting with antibodies for p53, Bax and β-actin. The p53 and Bax protein expression level was quantified densitometrically. A. Normal control group; B. Model control group; C. Matrine-treated group-L; D. Matrine-treated group-M; and E. Matrine-treated group-H

The dose-dependent effect of matrine on cleaved capase-3 expression. The cleaved capase-3 protein expression level was quantified densitometrically. A. Normal control group; B. Model control group; C. Matrine-treated group-L; D. Matrine-treated group-M; and E. Matrine-treated group-H

Matrine decreases MLCK expression. Immunohistochemical analyses of immuno- staining for MLCK-positive foci (400×) were performed on liver tissues from control, model and matrine-treated rats. A. Normal control group; B. Model control group; C. Matrine-treated group-L; D. Matrine-treated group-M; and E. Matrine-treated group-H. **P < 0.01 vs the control group; ▲P < 0.05 and ▲▲P < 0.01 vs the model group
