Animals
Forty eight adult male albino Wistar rats (Pasteur Institute, Tehran, Iran) weighing 200–250 g were used in these experiments. The rats were acclimated to the vivarium (a 12 h light/dark cycle at a temperature controlled room, 23 ± 1 °C), for at least one week prior to the onset of the experiments, with free access to chow and tap water. The animals were randomly allocated to different experimental groups. Each animal was used only once. All experiments were executed in accordance with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health Publication No.80-23, revised 1996) and were approved by the Research and Ethics Committee of Shahid Beheshti University of Medical Science.
Stereotaxic surgery
Rats were anesthetized with intraperitoneal (i.p.) injection of ketamine (100 mg/Kg) and xylazine (10 mg/Kg). Cannulae (23-gauge diameter, 11 mm long) were stereotaxically (stereotaxic apparatus, Stoelting, USA) implanted into the LH and NAc. The coordinates for these regions were determined by the rat brain atlas of Paxinos and Watson (2007) as AP = 2.6 ±0.15 mm caudal to the bregma, Lat = ±1.4 mm lateral to midline and DV = 8.5 mm ventral from the skull surface for the LH and for the NAc was AP = 1.6 ± 0.15 mm rostral to the bregma, Lat = ±1.6mm lateral to midline, DV = 7.8 mm ventral from the skull surface (
20). The guide cannulae were secured in place using two stainless steel screws anchored to the skull and dental acryl cement. After the cement was completely dried and hardened, two stainless steel stylets inserted into the each guide cannulae to prevent occlusion during the recovery period. Then, animals were housed randomly and allowed to recover for a period of 5–7 days before the experiments.
Drugs
All drugs were freshly prepared on the day of the experiment. Carbachol (Carbamoylcholine chloride; Tocris Bioscience, Bristol, UK) as a cholinergic agonist dissolved in physiological saline and SB334867 (Tocris Bioscience, Bristol, UK); as an OX1 receptor antagonist dissolved in 12% dimethyl sulfoxide (DMSO; Sigma–Aldrich, Germany). 2.5%formalin was prepared by diluting 37% formaldehyde (i.e., the commercially available saturated aqueous solution of formaldehyde, Merk, Germany) with physiological saline solution.
Drug administration
Microinjections were performed by lowering a stainless steel injector cannula (30-gauge needle) with a length of 1 mm longer than the guide cannulae into the LH and NAc. The injector cannula was attached to a 1-µL Hamilton syringe by polyethylene tubing (PE-20), subsequently the drug solution or vehicles was infused unilaterally over 60s and was left for an additional 60s to prevent the backflow of drugs. Carbachol or saline were administered in a total volume 0.5 µL into the LH and SB334867 or 12% DMSO were administered in a total volume of 0.5 µL into the NAc.
Formalin test
All rats were injected subcutaneously into the plantar surface of the right hind paw by formalin (2.5%; 50 µL), 5 min after microinjection of either the drugs or vehicles into the LH or NAc. Then placed in a 35 × 35 × 35 cm sized transparent plastic chamber with a mirror angled at 45˚ behind the box which used for observing the rat’s behavior during the test. Nociceptive behaviorswere observed in these experiments for 60-min period following formalin injection. During this time, pain scores were determined for every 5-min block by measuring the amount of time spent in each type of the four behavioral categories: 0, the position and posture of the injected hind paw was indistinguishable from another hind paw; 1, the injected paw had little or no weight on it; 2, the injected paw was elevated and did not have contact with any surface; 3, the injected paw was licked, bitten or shaken (
18,
21). Then, a weighted nociceptive score, with a range of 0-3, was calculated by multiplying the time spent in each category by the category weigh, summing these products and dividing by the total time (300 s) for each 5-min block of time:
Pain score = (t0 × 0) + (t1 × 1) + (t2 × 2) + (t3 × 3)/t0+t1+t2+t3
Formalin injection provokes a biphasic nociceptive response. The first 5-min block is considered as early phase (0–5 min), and the late phase takes more time and is defined from 15 to 60min after formalin injection into the hind paw. By utilizing this method, an ordinal scale (
18) of nociceptive scores was generated with a range of 0–3.
Experimental design
The dose–response effects of intra-LH administration of carbachol has been published in our laboratory`s previous work (
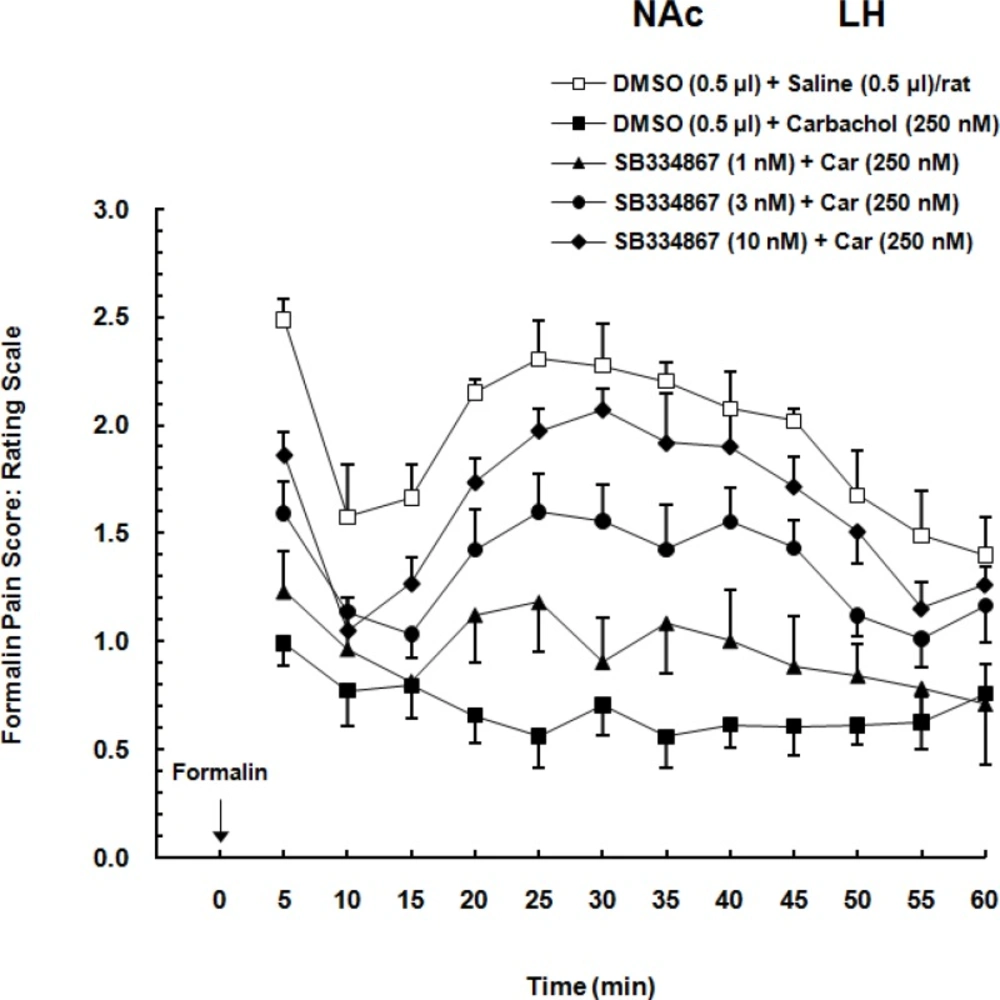
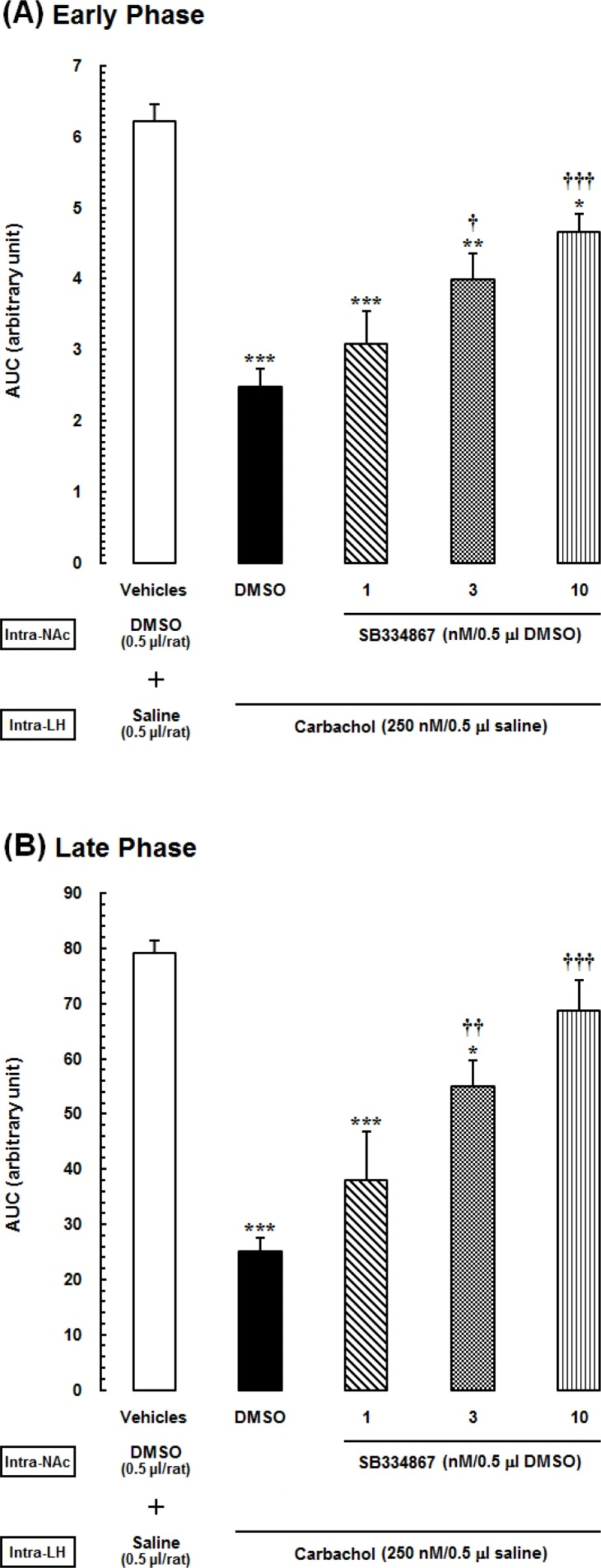
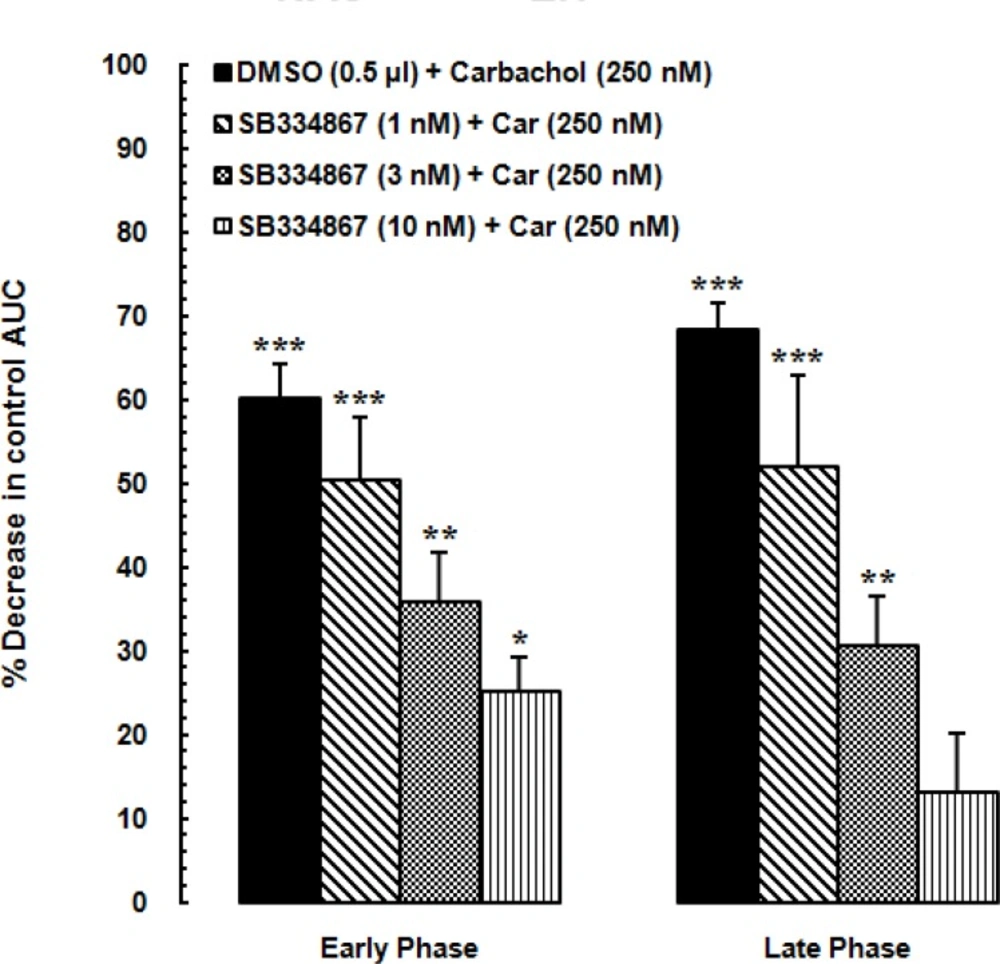
21). In that study carbachol microinjected to the LH in four doses (62.5, 125, 250 and 500 nM/0. 5 µL saline) 5 min before injection of formalin. The important result of the mentioned study was that intra-LH carbachol dose-dependently could block the nociceptive responses induced by formalin injection in both phases and the maximum effect was seen at the doses of the 250 and 500 nM. Accordingly, in this study, we used the dose of 250 nM.
Effect of intra-accumbal administration of OX1 receptor antagonist on antinociception induced by chemical stimulation of the LH
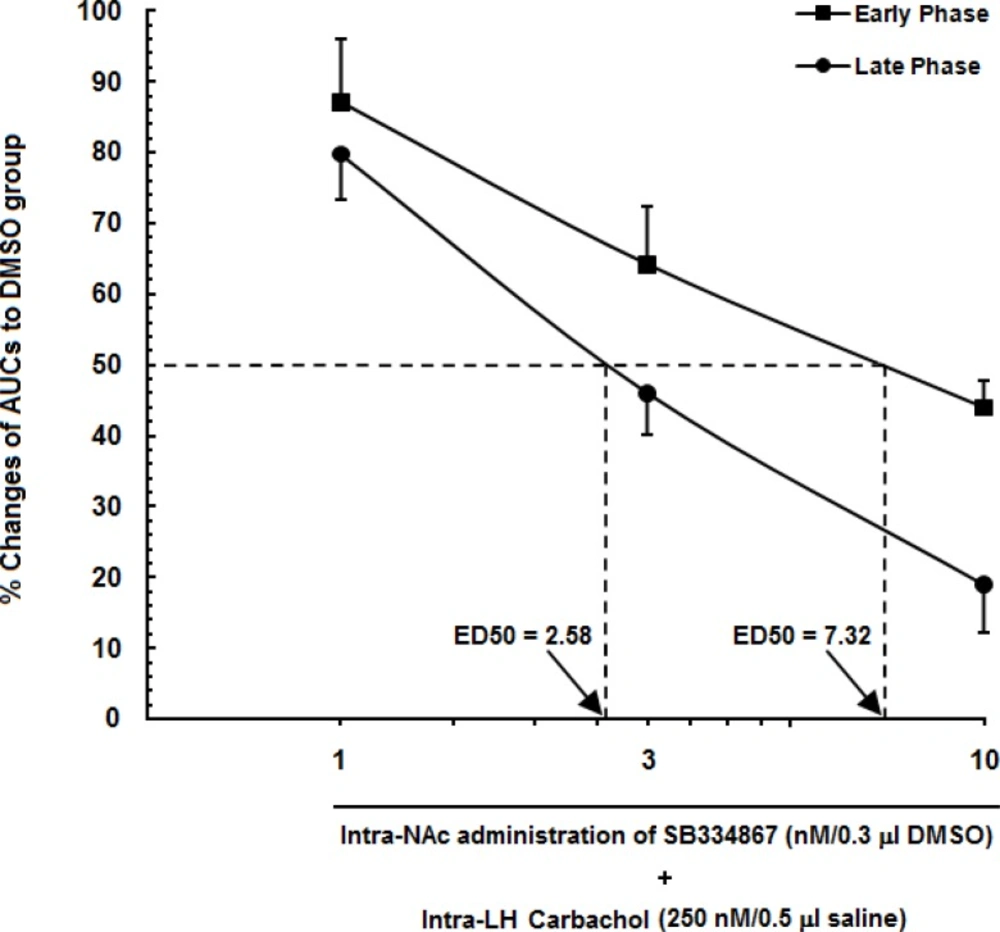
To examine the role of intra-accumbal OX1 receptors in the LH stimulation-induced antinociception, different doses of SB334867 (1, 3 and 10 nM/0.5µL DMSO) were unilaterally injected into the NAc, 5min prior to intra-LH administration of carbachol (effective dose; 250 nM/0.5 µL) and then test for formalin test. In the carbachol-control group rats received 12% DMSO instead of SB334867 and vehicles-control group, received 12% DMSO and saline (0.5 µL) into the NAc and LH, respectively.
Histological verification
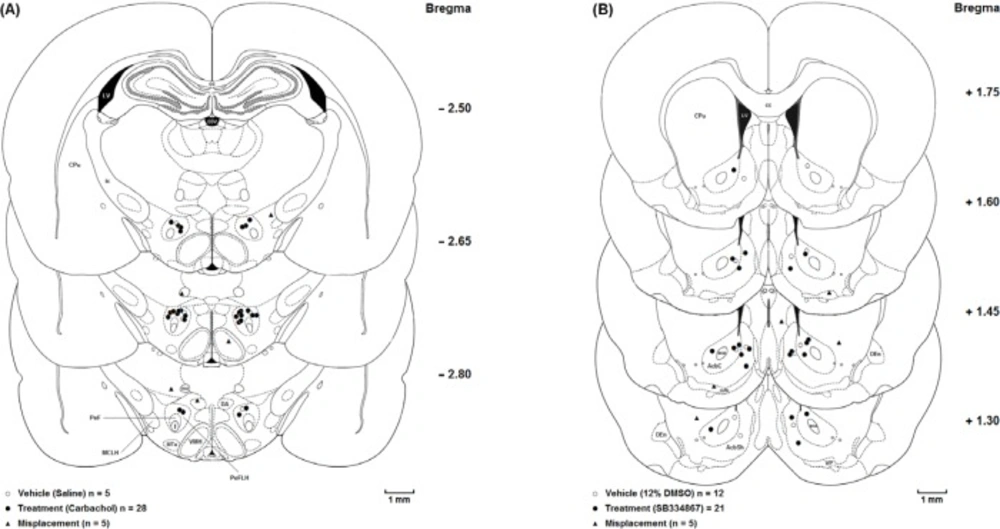
The animals were deeply anesthetized with ketamine and xylazine (100 mg/Kg and 10 mg/Kg, respectively) after the experiments were accomplished. Transcardial perfusion was carried out with 0.9% saline and 10% formalin solution. The brains were removed and cut coronally in 50-µM sections through the cannulae placements. The neuroanatomical locations of cannulae tips were confirmed using rat brain atlas (
20). Only the results from animals with correct cannulae placements were included in the data analysis (
Figure 1.).
Statistics
The obtained data are expressed as mean ± SEM (standard error of mean). To specify the treatment and time effects on the pain behaviors, the mean nociceptive scores of different groups were compared by two-way ANOVA followed by Bonferroni’s multiple comparisons test. In order to evaluate the nociceptive responses, area under the curve (AUC) was calculated as raw pain scores × time by linear trapezoidal method and a single value was used in statistical analyses. We calculated the AUC of the early phase (1-5 min) and late phase (15-60 min) separately. The calculated AUC values in all experimental groups were subjected to one-way ANOVA followed by protected Newman–Keuls test for multiple comparisons. One-way ANOVA followed by Dunnett›s test was used to calculate the percentage decrease of the AUC values calculated for pain scores in the carbachol-control and experimental groups compared to AUC values of vehicle-control group during the early and late phases. Finally, two-way ANOVA followed by Bonferroni›s test was used to determine the effect of time (early or late phase of formalin-induced pain) and treatment on the percentage changes of AUCs of treatment groups compared to the AUC values of saline group. P-values less than 0.05 were considered to be statistically significant.