Chemicals and reagents
Linarin (>98% purity, isolated from Cirsium setosum) was obtained from Chengdu Must Bio-technology Co. Ltd (Chengdu, China). Huperzine A was obtained from National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). Acetylthiocholine iodide (ATCI) and 5,5'-dithiobis-(2-nitrobenzoic acid)(DTNB) were obtained from Shanghai Ruji Bio-technology Co. Ltd (Shanghai, China). Dimethyl sulfoxide (DMSO) was purchased from Tianjin Bodi Chemical Co. Ltd (Tianjin, Chian). All other reagents and organic solvents were of analytical grade.
Animals
Healthy male Kunming (KM) mice, weighing 20 ± 2 g, SPF, were provided by the Laboratory Animal Center of Shenyang Pharmaceutical University (Shenyang, China). All animals were housed under standard controlled environmental conditions: temperature (25 ± 2 °C), relative humidity (60 ± 10%), room air change (12–18 times/h) and light/dark cycle (12/12). Food and water were available ad libitum. The study was conducted under protocols approved by the Animal Ethics Committee of Shenyang Pharmaceutical University, in accordance with the Guide for the Care and Use of Laboratory Animals (NIH publication No.85-23, revised in 1985).
In-vitro AChE inhibition assay
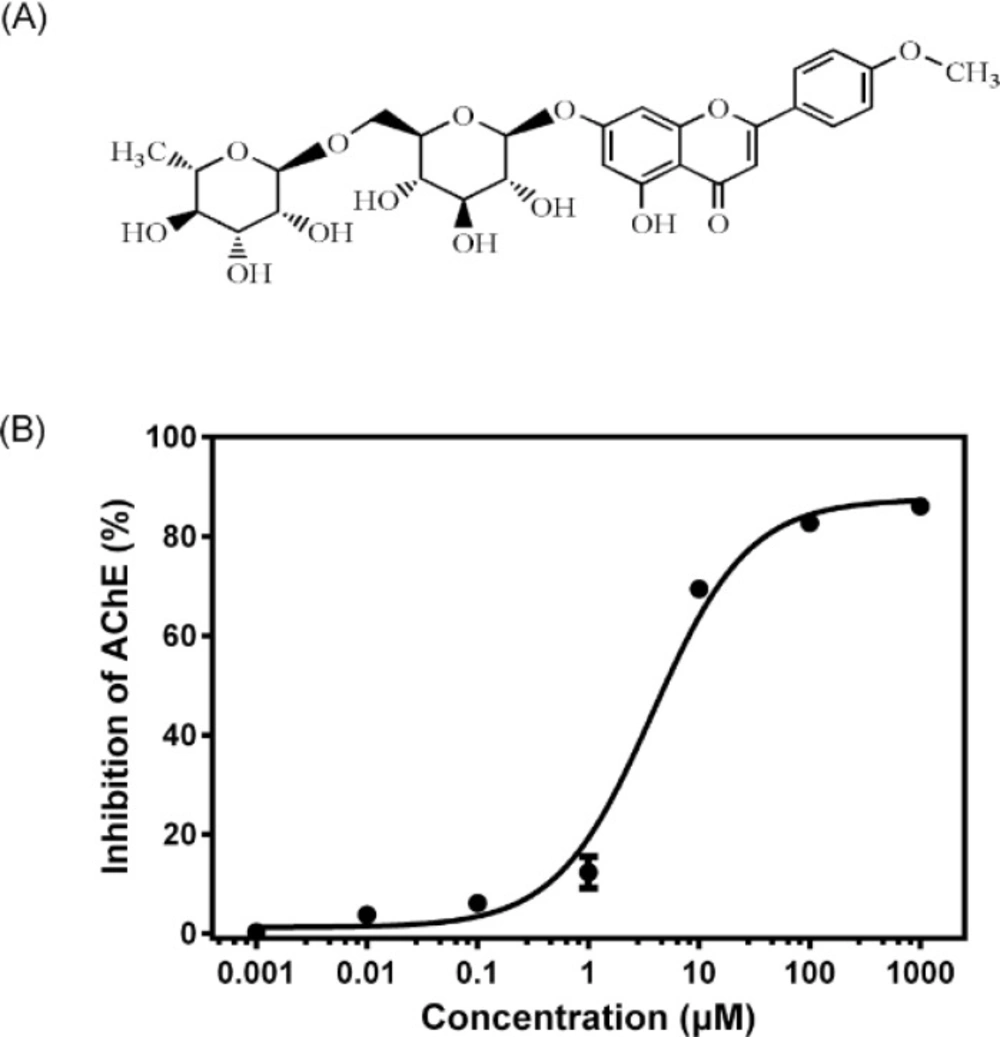
After the mice were killed by decapitation, the brains were quickly removed on an ice-cold plate and homogenized in a 10-fold volume of cold 10 mM phosphate buffer (pH 7.4). The homogenates were centrifuged at 3500 rpm for 10 min and the supernatants were employed as sources of enzyme. All the above steps were carried out at 4 °C. AChE activity was determined according to a modified Ellman assay method (
14). Briefly, 150 μL of the brain homogenates and 600 μL of different concentrations (0.001–1000 μM) of linarin (dissolved in DMSO and then diluted in 10 mM phosphate buffer with final DMSO concentration less than 1%) were mixed. After the mixture was pre-incubated at37°C for 5 min, 2100 μL of 1 mM DTNB and 300 μL of 3 mM ATCI in 10 mM phosphate buffer (pH 7.4) were added to start the reaction. The increase in absorbance due to the formation of 5-thio-2-nitrobenzoate anion was measured during 1 min at 405 nm and 37 °C. Huperzine A at a concentration of 150 μM was used as a positive control. The AChE activity was estimated through changes of absorbance per minute and the results were reported as percentage of inhibition to AChE activity, where the optical density measured from blank (without inhibitor) was considered to be 0% of inhibition and the optical density measured from control (huperzine A) was considered to be 100%. The percentage of inhibition was calculated as follows: (
A blank −
A sample)/(
A blank −
A control)× 100%, where
A blank,
A sample and
A control are the absorbance of blank, sample and positive control, respectively. The half maximal inhibitory concentration (IC
50) was calculated from the Prism (GraphPad 6.0, Dose-response - Inhibition) curve obtained by plotting the percentage inhibition versus the concentration of linarin.
Ex-vivo AChE inhibition assay
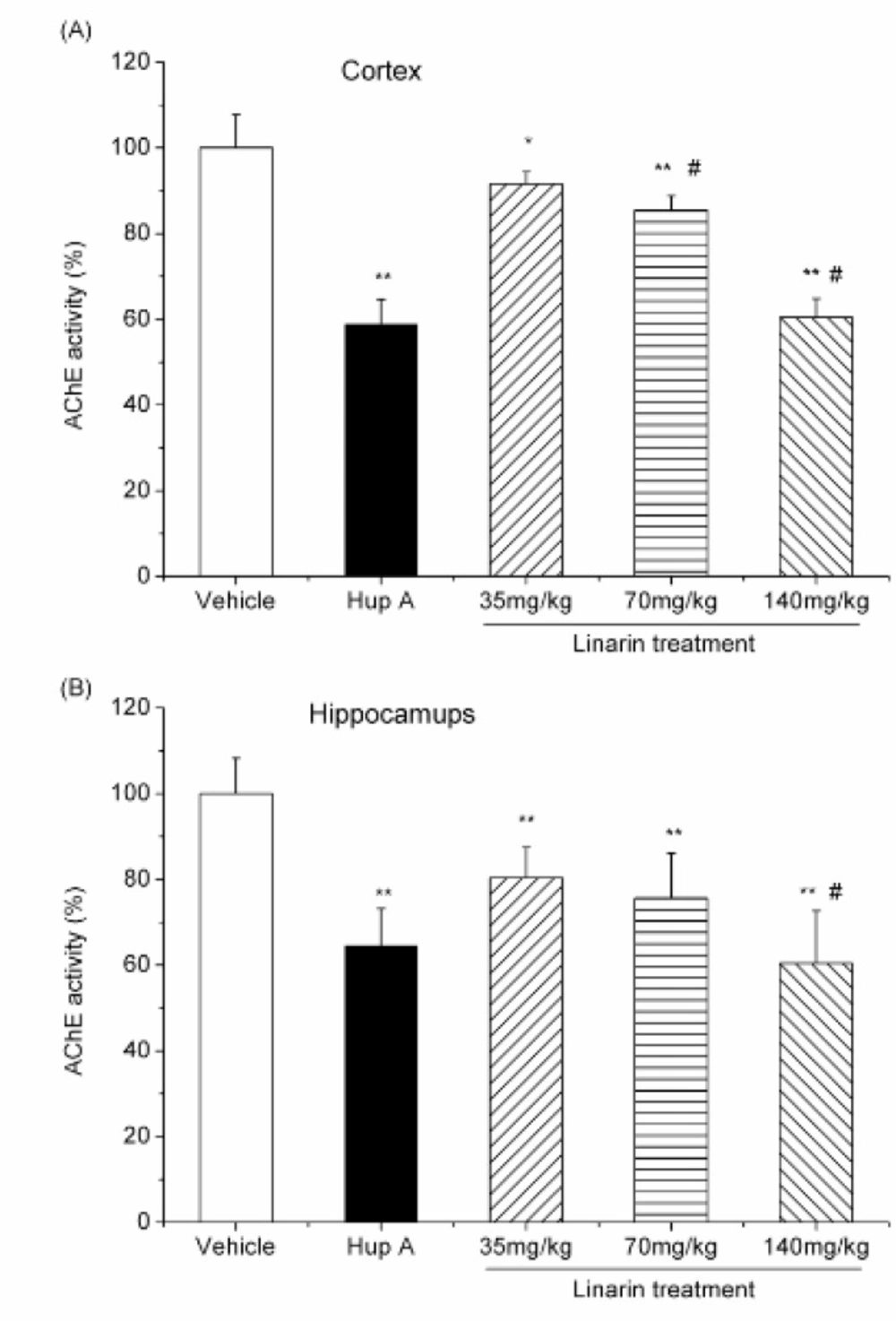
The mice were randomly divided into five groups of six mice each: normal control group, huperzine A (0.5 mg/Kg) treated group and low-, moderate-, high-dose linarin(35, 70 and 140 mg/Kg) treated groups. The mice in the normal control group received an equivalent volume of 5% DMSO in saline (vehicle). All the mice were treated with a single intraperitoneal injection of the respective drugs and sacrificed 30 min later. The brains were quickly removed on an ice-cold plate, and then the cortex and hippocampi were dissected out. These tissues were homogenized in a 10-fold volume of cold 10 mM phosphate buffer (pH 7.4). The homogenates were centrifuged at 3500 rpm for 10 min and the supernatants were employed as sources of enzyme in AChE assay as described above. The percentage of inhibition was calculated by comparison with the AChE activity of mice treated with vehicle.
Statistical analyses
Statistical analyses were performed by using one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test using the Statistical Package for the Social Sciences (SPSS) software.
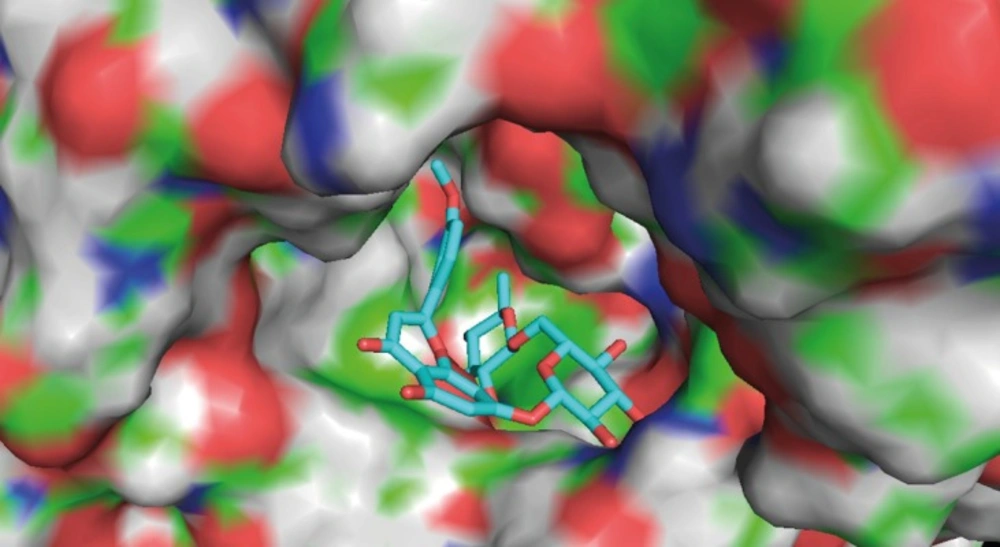
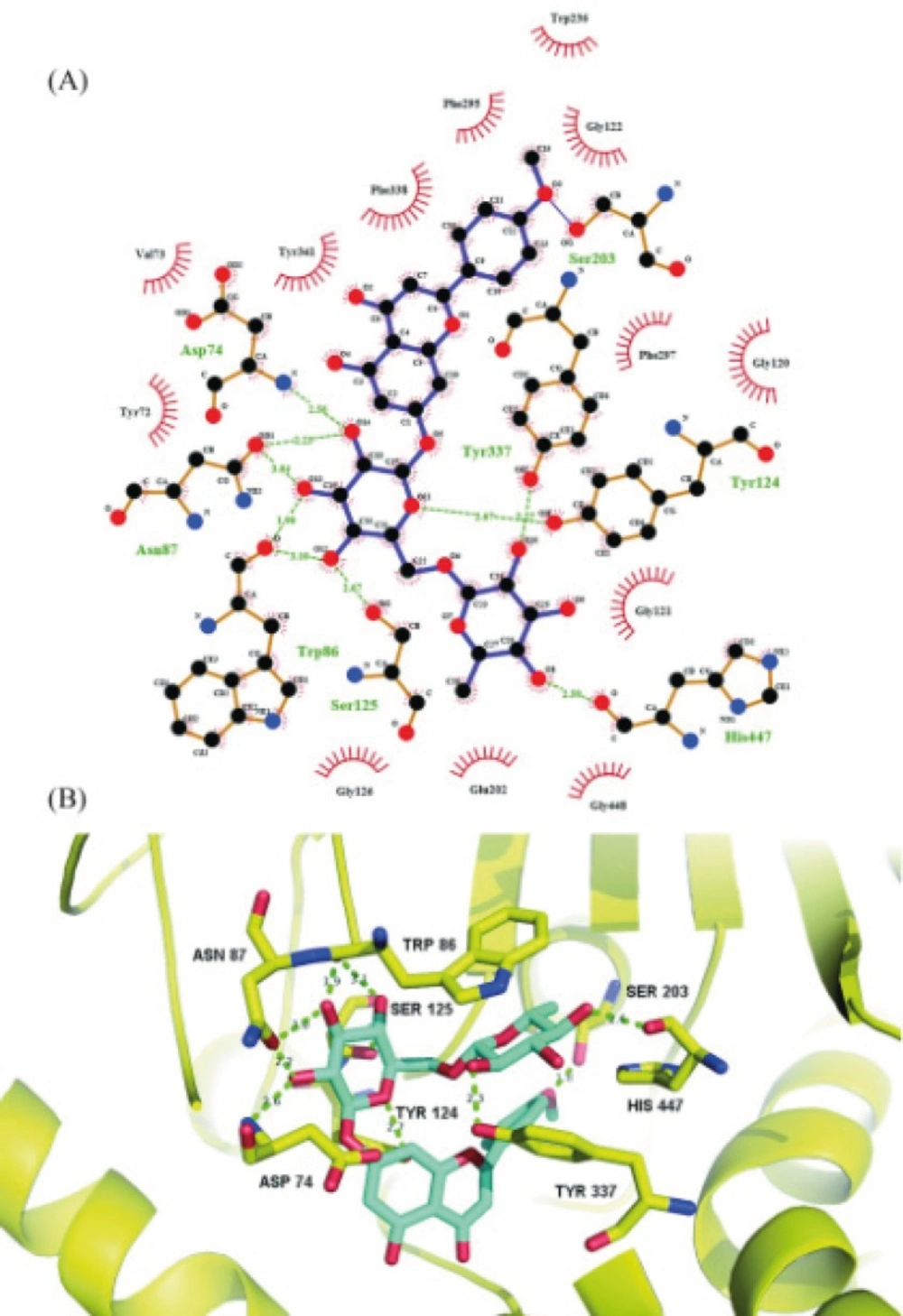
Molecular docking simulation
Molecular docking simulation was performed using AutoDock 4.2. The X-ray crystal structure of acetylcholinesterase was obtained from RCSB protein data bank, which was prepared by removing both non-polar hydrogens and water molecules, and adding Kollman united-atom partial charges and solvation parameters. Meanwhile, the torsion angles of linarin were identified. A grid box of 70×70×70 points in the x, y and z directions were constructed with a grid spacing of 0.375 Å at the center of the catalytic pocket of acetylcholinesterase. The Lamarckian Genetic Algorithm method was used for the global search for optimum binding position. After completion of docking and clustering analysis, the most favorable binding conformation with the lowest free energies was selected as the binding pose.