Construction of expression vector
One of the most important elements for transporting a recombinant protein into the periplasmic space is an N-terminal signal sequence (
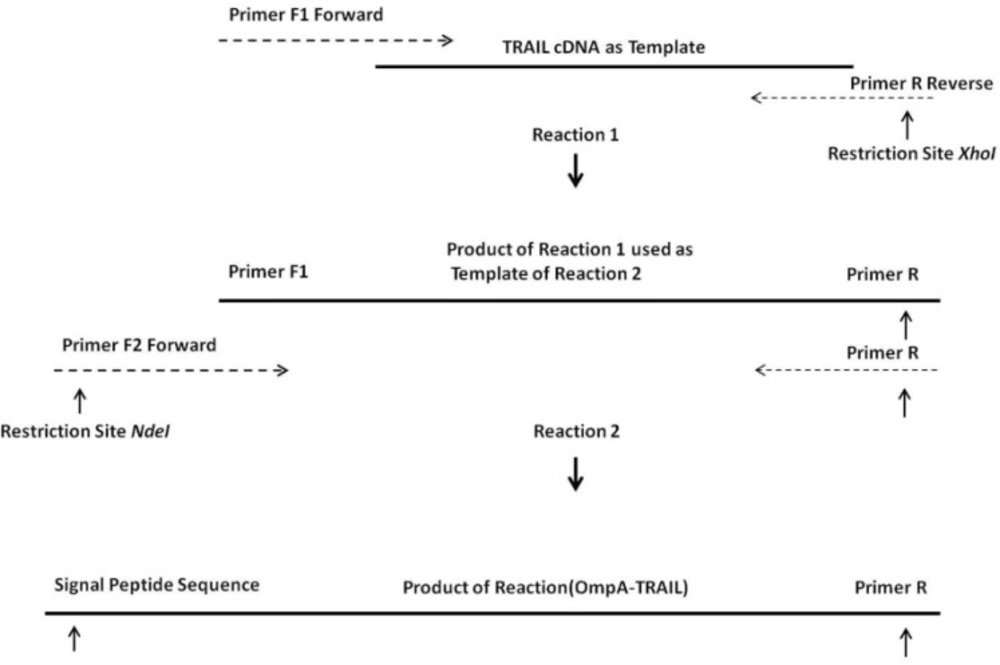
24). For this purpose, an OmpA signal sequence was fused to the 5´ region of the TRAIL cDNA using two sequential OE-PCR reactions (
Figure 1). The OmpA signal sequence was chosen because it functions efficiently to secrete a large amount of the OmpA protein (
25) and it is frequently used in several studies attempting to secrete different proteins (-). As expected, the size of the PCR products sequentially increased from 504 bp to 594 bp indicating a correct fusion of the OmpA signal sequence to TRAIL cDNA (
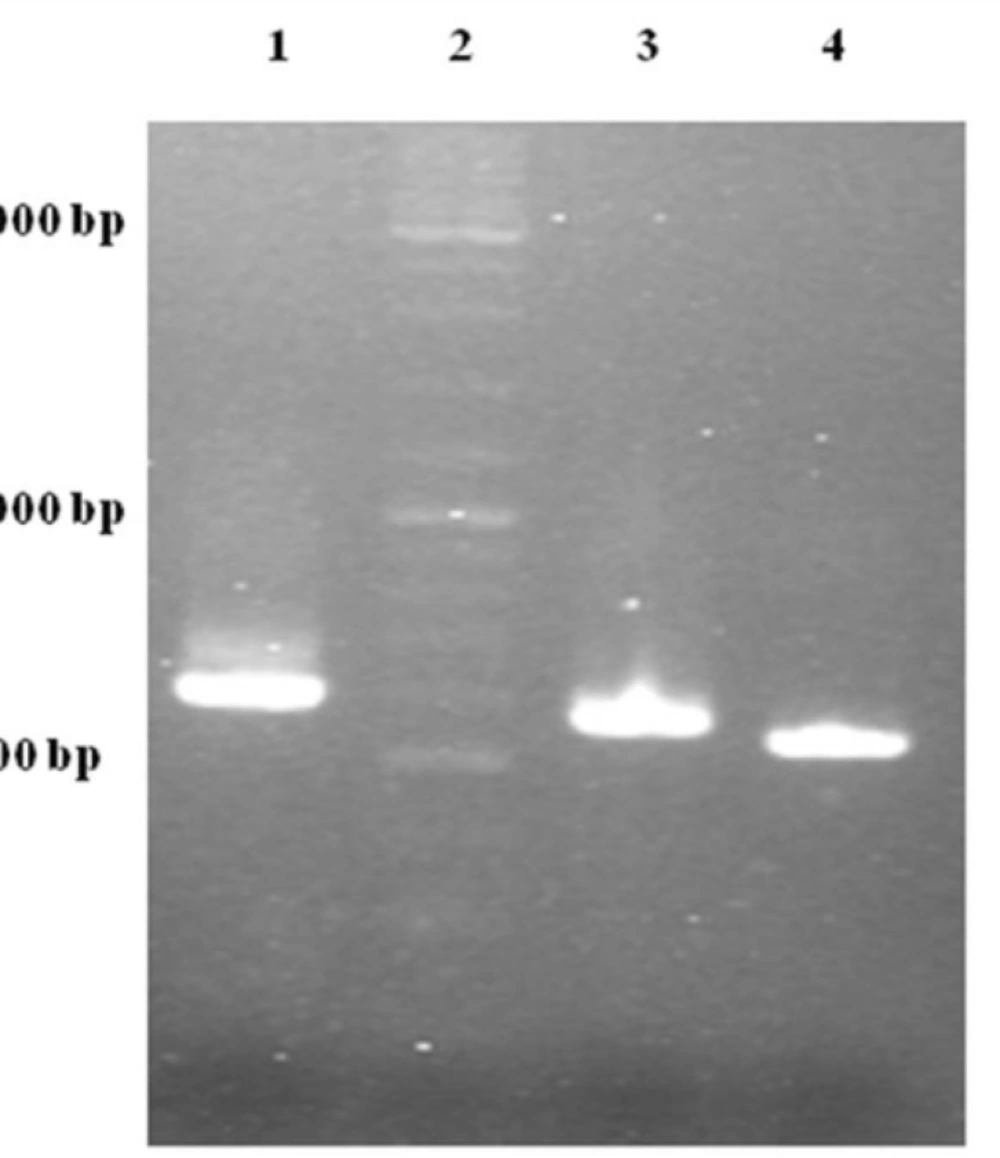
Figure 3).
The increase in size of the products after each PCR reaction demonstrates that the 5´ region of the TRAIL fragment is extended due to the primer design (
Table 1). In the first cycles of each PCR reaction, the forward primer was hybridized with 5´ region of template by 19-25 overlapping nucleotides of its own 3´ region. Then, in the later cycles, the overhang sequence of the forward primer was inserted into the template gradually. Therefore, in the first 5 cycles, annealing temperature was considered 65°C according to the overlapping region, whereas in the next 30 cycles it was considered 69°C according to the whole primer sequence. In addition, a high-fidelity DNA polymerase pfu was used to ensure that the rate of mutations in sequential PCR reactions was minimal.
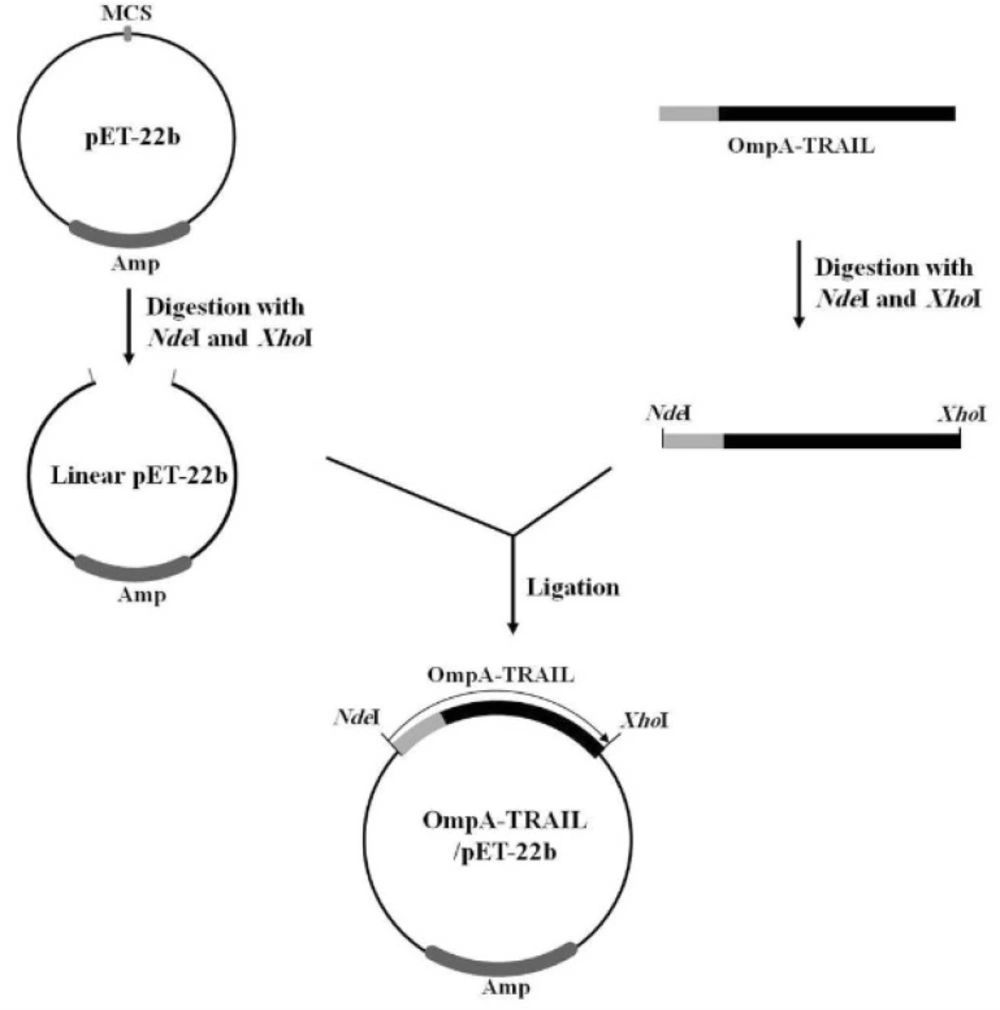
Finally, in order to express recombinant TRAIL, the PCR product was purified, digested with NdeI and XhoI, and then sub-cloned into the corresponding sites of the pET-22b plasmid. The correct insertion of OmpA-TRAIL into the recombinant plasmid was confirmed by PCR analysis and DNA sequencing.
Gel electrophoresis analysis of OE-PCR products. Lane 1: 594 bp product amplified with F2R primer indicating OmpA-TRAIL whole sequence. Lane 2: DNA Ladder marker. Lane 3: 557 bp product amplified with F1R primer indicating partially-synthesized signal sequence. Lane 4: 504 bp product amplified with TRAIL specific primers indicating TRAIL sequence.
Expression and purification of recombinant TRAIL
The ability of recombinant plasmid OmpA-TRAIL/pET22b to express and transport the recombinant protein into the periplasmic space was analyzed by expression induction and then isolation of periplasmic and cytoplasmic proteins. The recombinant plasmid was transformed into the E.coli BL21 (DE3) and cultivated in TB culture medium followed by induction of TRAIL expression with 0.1 mM IPTG. In order to release periplasmic proteins, outer membrane was selectively disrupted by osmotic shock procedure. Cytoplasmic proteins were then released by disruption of resulted spheroplasts. Whole cells were also disrupted to assess the total expression of TRAIL.
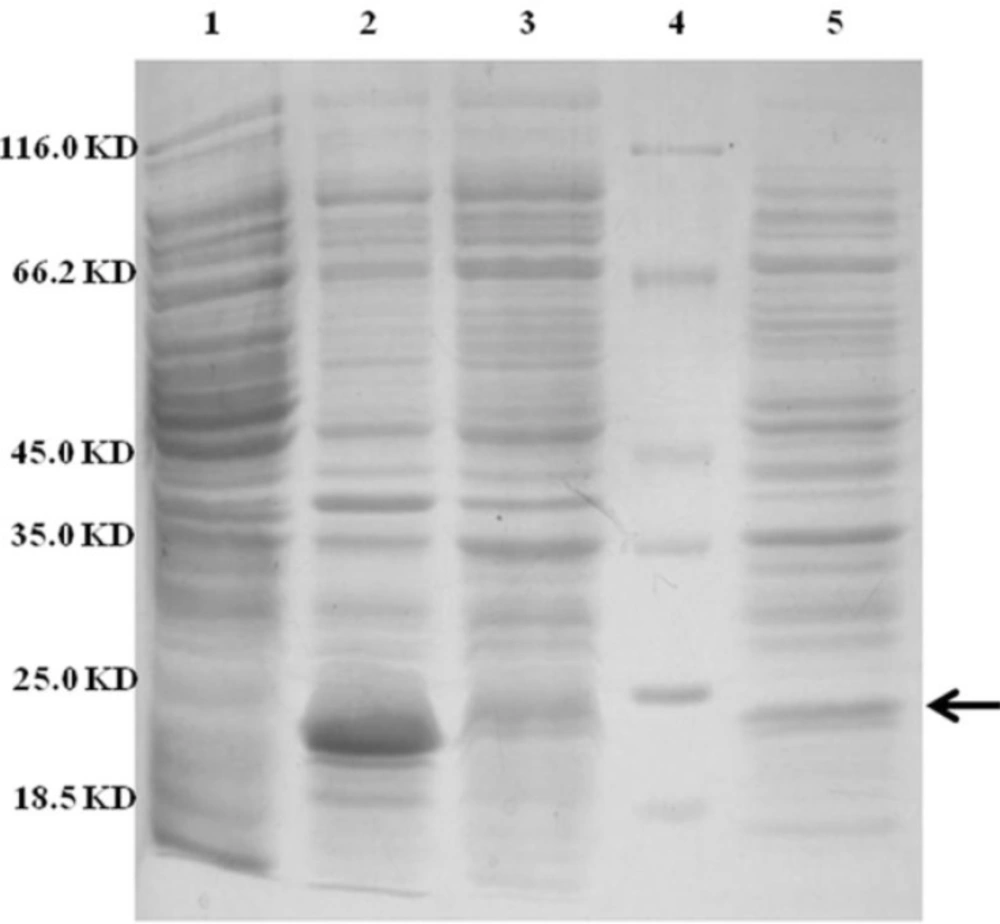
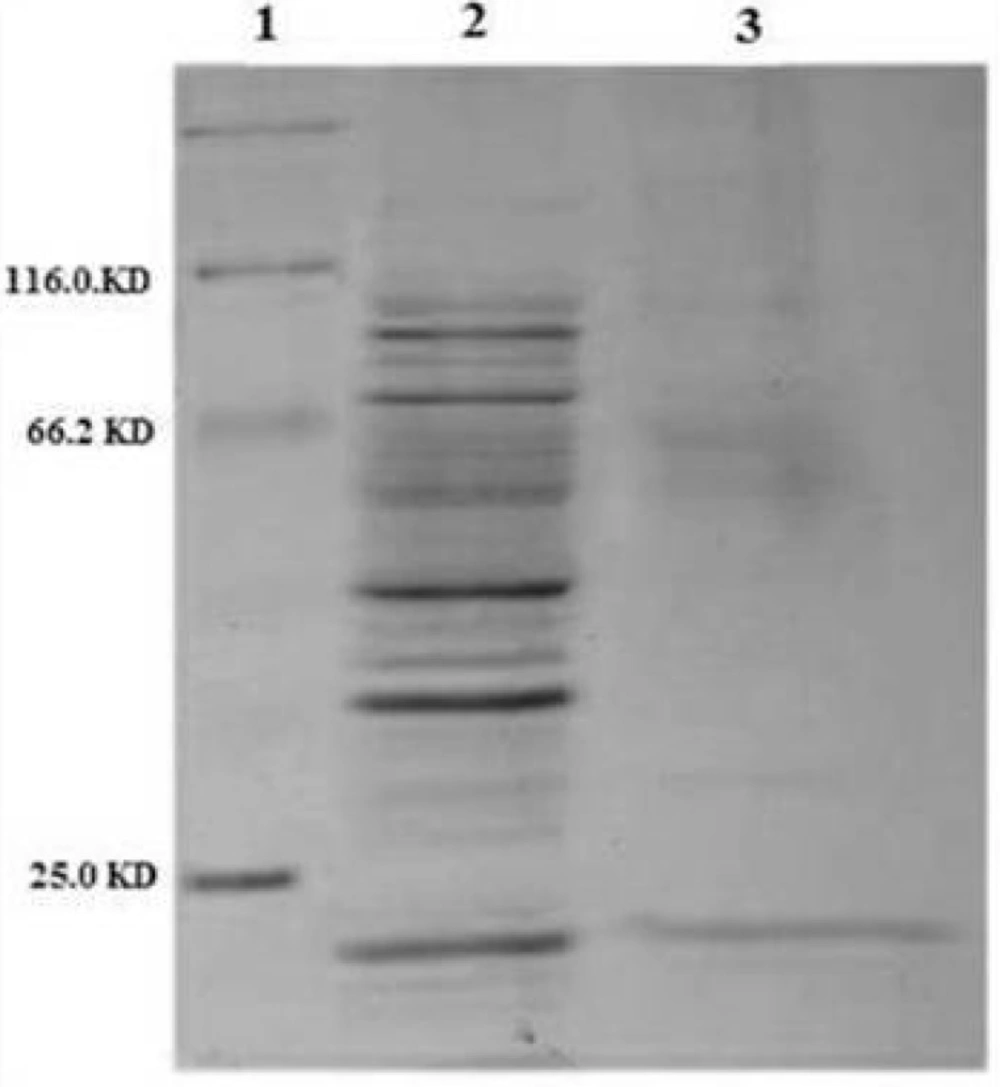
Approximately 37% of total expressed TRAIL was secreted into the periplasmic space based on measuring the staining intensity of protein bands using ImageJ software (NIH) (
Figure 4). The results are consistent with the recent report of Sockolosky
et al. (
26) showing that the recombinant human growth hormone is directed to the
E. coli periplasm using the pET based expression based on OmpA signal sequence. Also, several studies have shown the efficacy of the OmpA signal sequence to direct recombinant proteins into the periplasmic space (
29,
30)
SDS-PAGE analysis of different cellular fractions of E.coli BL21(DE3) including OmpA-TRAIL. Lane1 and 2: whole cell before and after induction, respectively. Lane 3: cytoplasmic fraction after induction. Lane 4: protein marker. Lane 5: periplasmic fraction after induction. 10 µg of proteins were loaded in each lane.
However, although the level of TRAIL secretion into periplasm was satisfactory, it seems that a considerable amount of recombinant TRAIL was remained in
E.coli cytoplasm. It might be explained by a rather high expression rate and, therefore, lack of time for interaction between the signal sequence and the transport pathway agents leading to remaining target protein in the cytoplasm as inclusion bodies. (
31). Low concentration of IPTG (0.1 mM) was applied in order to control the expression, but high cultivation temperature (37 ºC) may be responsible for high expression rate. (
16,
32). On the other hand, secretion into the periplasm is a complex process and the presence of a signal sequence does not always ensure the efficient protein secretion (
18).
Subsequently, periplasmic TRAIL was purified for further study on its biological activity. Cloning in vector pET-22b fuses a histag to the C-terminal of expressed protein which was used for purification of periplasmic TRAIL by Ni-NTA affinity chromatography.
Figure 5 shows an efficient purification of the periplasmic TRAIL in which other contaminating proteins were mostly removed. The obtained results of purification study were summarized in
Table 2.
Eluting buffer has some components such as imidazole which interfere with the assessment of biological activity of TRAIL (
33). For this reason, eluting fractions were dialyzed against PBS buffer to remove any interfering components.
SDS-PAGE analysis of purified periplasmic TRAIL by Ni-NTA affinity chromatography. Lane 1: protein marker. Lane 2: 10 µg of periplasmic fraction of E.coli BL21 (DE3) after induction. Lane 3: 1 µg of purified TRAIL after elution by imidazole
| Sample | Volume (mL) | Total Periplasmic Protein (mg)a | Periplasmic TRAIL (mg) b | Purity (%) | Purification Yield (%) |
|---|
| Culture | 100 | 5 | 0.78 | 16 | - |
| Purified Solution | 1 | 0.75 | 0.68 | 91 | 87 |
Protein concentration was determined based on absorbance at 280 nm.
TRAIL concentration and purity were estimated by scanning the SDS-PAGE gel using ImageJ software.
Immunological identification of recombinant TRAIL
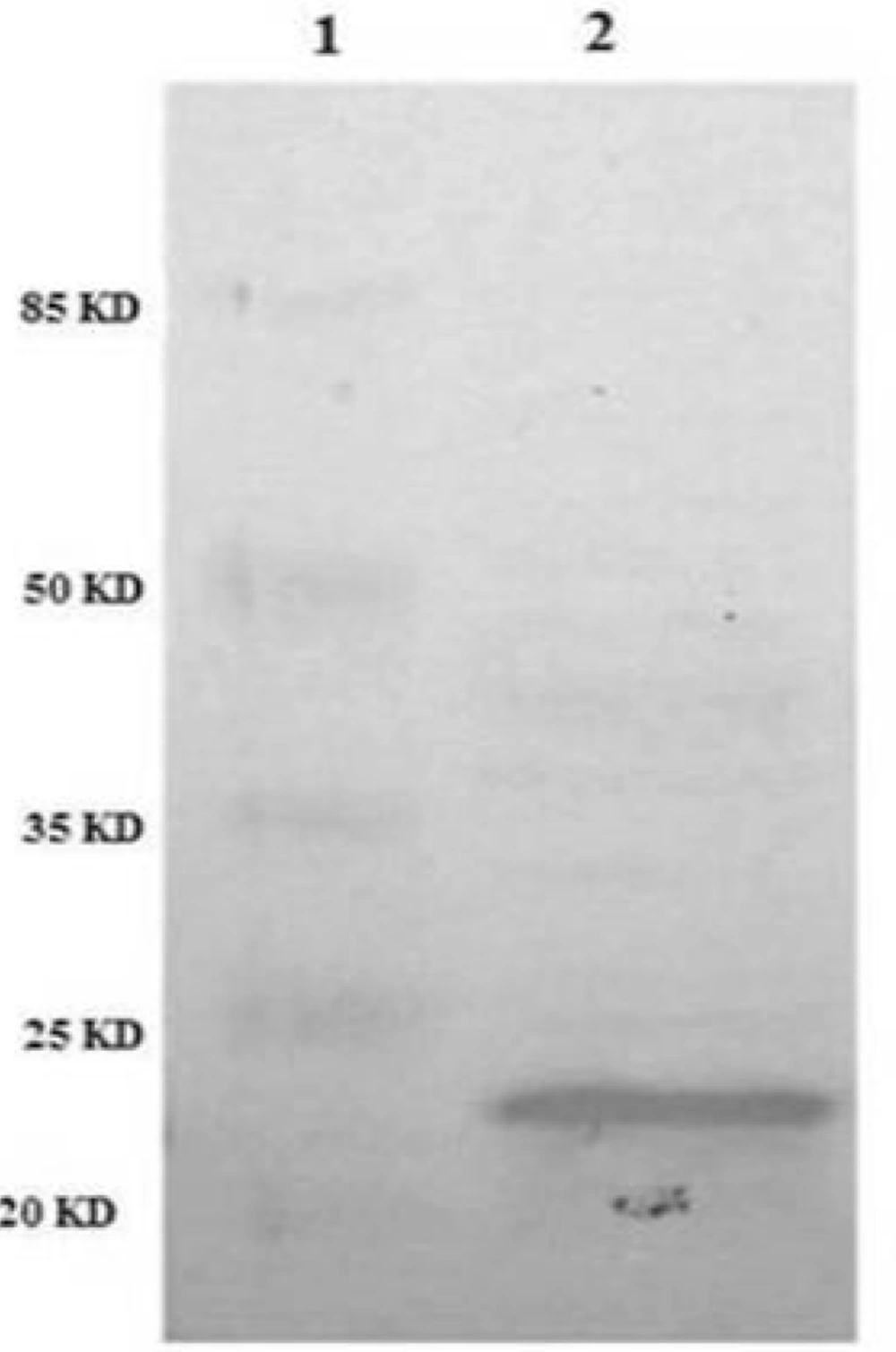
In order to confirm the identity of recombinant TRAIL immunologically, western blot analysis was performed using anti-TRAIL antibody. The results showed that periplasmic fraction reacted with anti-TRAIL antibody and the size of the protein was approximately 21 kD as expected for recombinant TRAIL (
Figure 6).
Identification of periplasmic TRAIL protein by Western Blot. TRAIL was resolved by SDS-PAGE and then transferred to nitrocellulose membrane. After incubation with anti-TRAIL antibody and then secondary antibody, the membrane was developed with the BCIP/NBT substrate system. Lane 1: protein marker. Lane 2: periplasmic TRAIL.
Biological activity of TRAIL
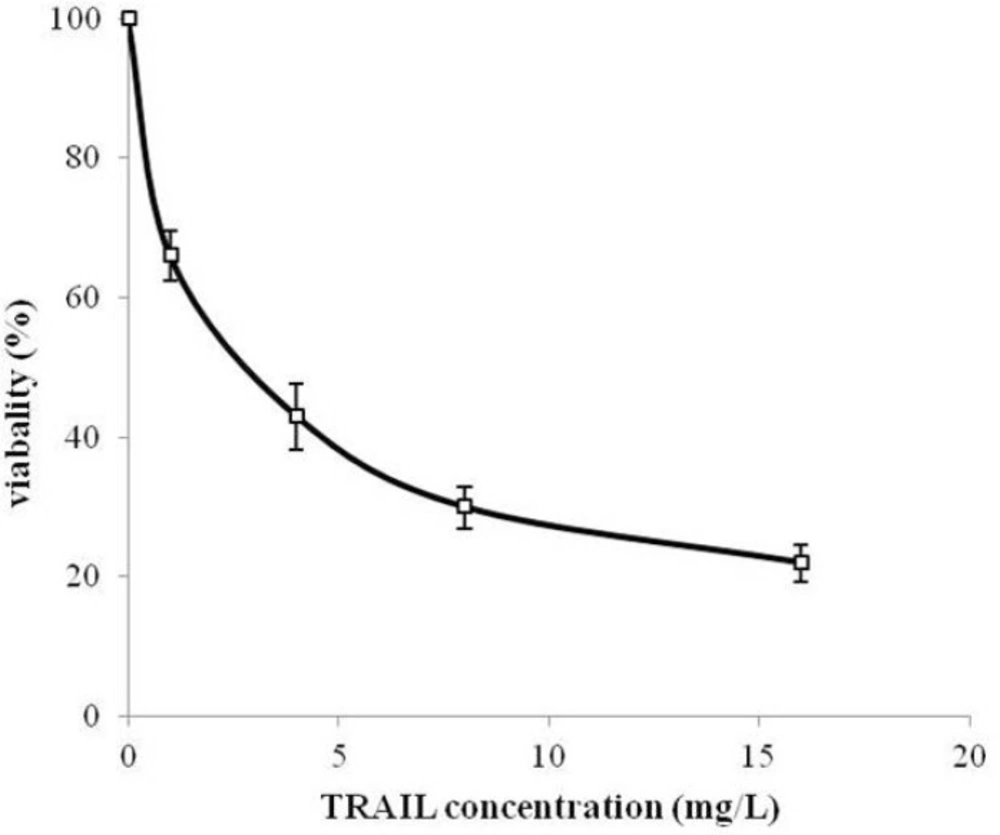
MTT assay protocol was performed to evaluate the cytotoxicity effect of periplasmic expressed TRAIL against human cervical cancer HeLa cells. HeLa is one of the most frequently used cell lines in pharmaceutical research studies for assessment the anti-tumor activity (
34,
35). Periplasmic expressed TRAIL induced cytotoxicity in HeLa cells in a dose-dependent manner (
Figure 7). The cytotoxicity was detectable for concentration of 200 µg/L of periplasmic TRAIL. Also, the ED
50 was about 2.7 mg/L determined by GraphPad Prism software (version 6.0, Graphpad software). This cytotoxicity effect was in accordance with the previous studies (
7,
31). However, in those studies, recombinant TRAIL was expressed as inclusion body in
E.coli and needed to be refolded before assessing its cytotoxic effect, while, in the present study, periplasmic TRAIL was soluble and biologically active. Therefore, it could be concluded that TRAIL directed to the periplasm has a proper folding. In other expression systems (
e.g.
P.pastoris), recombinant TRAIL has shown cytotoxic effect against various cancer cell lines (-). However, it seems that efficiency of TRAIL cytotoxic effect may partially be varied depending on cancer cell lines, expression systems, and source of recombinant TRAIL (
i.e. human or animal) (
36).
In this paper a simple method for production of the recombinant TRAIL directed to the E. coli periplasm is presented using pET expression system and OmpA signal sequence. The results demonstrated efficient periplasmic expression of TRAIL. Considering the advantages of directing recombinant TRAIL into the E.coli periplasmic space, this study may be useful for production of this therapeutic ligand in E.coli periplasm.
Cytotoxicity of periplasmic expressed TRAIL in human cervical cancer HeLa cells. HeLa cells were dispensed into 96-well plate (2×103 cells/well) and treated with different concentrations of TRAIL for 24 h. Cytotoxicity was then assessed based on MTT assay protocol. All measurements are reported as mean ± SD (n=3).